
Research | DOI: https://doi.org/10.31579/2637-8914/055
*Corresponding Author: Hamouda Elabed, Laboratory of Contagious Diseases and Biologically Active Substances LR99-ES27, Faculty of Pharmacy of Monastir, University of Monastir, Tunisia.
Citation: Rihab Ben Slama, Hamouda Elabed, Hanene Miladi, Tarek Zmantar, Maha Mastouri, et al. (2021). Morphological Alterations and Virulence Gene Expression of Listeria Monocytogenes Cells in Response to Gamma Irradiation Treatments. J. Nutrition and Food Processing, 4(5); DOI:10.31579/2637-8914/055
Copyright: 2021, Hamouda Elabed. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 10 June 2021 | Accepted: 30 June 2021 | Published: 09 July 2021
Keywords: : gamma irradiation; listeria monocytogenes; gene expression; fatty acid; AFM; agro-food industries; dehydrated vegetables; IAEA; food safety; potential pathogenic bacteria; chromatographic method
Gamma irradiation is one of the most popular ray treatments in food industry. It's used to control micro-organisms proliferation in a wide range of products however the response of bacteria to low doses is still unknown. In this study we mainly focus on morphological alteration and virulence gene expression in Listeria monocytogenes after gamma irradiation treatments. The atomic force micrographs (AFM) showed that 0.5 kGy dose has no effect on the membrane morphology of L. monocytogenes. However, after 0.7 kGy treatment, the cells lost their typical shape and smooth membrane and 1 kGy dose was totally destructive. Moreover, membrane fatty acid composition was analyzed by the chromatographic method after different gamma doses. Significant modifications on fatty acids composition were detected in the irradiated strain showing a novo synthesis of membrane lipids: C12:0; C14:0; C15:0; C16:0 and C18:0. In addition, we reported an increase of the saturated fatty acid, essential for membrane adaptation under stress conditions. The expression levels of three virulence genes (hlyA, fri and prfA) were studied in the same conditions using real-time PCR technique. The analysis revealed that both prfA and fri genes were up-regulated after gamma treatment. The induction of prfA, which is a regulator gene, may affect the expression other genes controlling the adaptive form in the treated strain. This study open prospects for further researches to explain the regulatory mechanisms of the adaptive response in Listeria monocytogenes when exposed to sublethal irradiation-stress.
In the Agro-Food Industries, the irradiation is the process of applying ionizing energy to foodstuffs to improve microbiological safety and to extend the shelf life [1]. Irradiation technology is used for decontamination and/or sterilization of dehydrated vegetables, fruits, meats, poultry, fish and seafood in order to improve product safety and shelf life [2, 3]. According to the International Atomic Energy Agency (IAEA), the quantities of foods that are irradiated are increasing each year and the majority of these foods are treated by gamma irradiation. Moreover, it is expected that more food will be treated by irradiation in the future, because this technology can address food safety and security problems without significantly affecting at the nutritional attributes [4].
The treatment depended on the process objective with respect radiological safety and generally, the doses used in gamma irradiation treatment are between 0.3 and 3 kGy [5-7]. For microbial decontamination, the targets of gamma irradiation are chromosomal DNA, RNA, cytoplasmic membrane and proteins [8, 9]. Escherichia coli, Toxoplasma gonkii, Salmonella enterica and Listeria monocytogenes were controlled in food using irradiation [10]. It has been reported that the Gram-positive bacteria is more resistant to gamma irradiation than other potential pathogenic bacteria like E. coli [11].
Listeria monocytogenes is a Gram-positive bacteria, frequently associated with food borne disease outbreaks. It is also considered as a major food safety challenge in the food industry. Improved control measures and risks assessments have greatly reduced the prevalence of this pathogen in food. However, the rate of illness has remained constant during the last decade [12]. The psychotropic nature of L. monocytogenes allows replication in refrigerated ready-to-eat food products that were contaminated during processing and packaging. Consequently, L. monocytogenes is characterized by widespread distribution and relatively high mortality rates [13]. The effectiveness of gamma irradiation in inactivating the L. monocytogens on different food products was previously reported; Song et al. confirmed the efficacy of gamma irradiation to inactivate foodborne pathogens on nuts indicating that the D-value of L. monocytogenes on pistachios was 1.02 kGy, larger than those of S. enterica or E. coli [14]. Other study has shown that L. monocytogenes and S. enterica growth was inactivated in fresh orange juice at 0-4 kGy [15]. Moreover, Niemira et al. evaluated the effect of gamma irradiation (0.3 and 0.6 kGy) on L. monocytogens inoculated cut pieces of endive, thereby demonstrating that irradiation and modified atmosphere packaging can be combined to prevent the regrowth of L. monocytogenes during post-irradiation refrigerated storage [16].
Listeria treatments using gamma rays have been widely applied for different goals; however, none of this research had studied the problems related to radio-resistant L. monocytogenes cells. Therefore, a better understanding of resistant cells is required to reduce listeriosis incidence. Adaptive response, morphological alteration and gene regulation should be key elements to study bacteria behavior. One of the most important issues related to the microbiological safety of irradiated foods is to know if the sub-lethal doses would make surviving bacterial cells more pathogenic.
The aim of this work was to study the responses elicited in Listeria monocytogenes following different gamma irradiation doses, by analyzing: (i) The morphological alteration using atomic force microscopy (ii) The membrane fatty acid profiles and (iii) The expression levels of three virulence genes.
1. Strains and culture conditions
The American Type Culture Collection Listeria monocytogenes ATCC 19115, serotype 4b [17] was used in this study. Cells were grown in tryptic soy broth with 0.6% yeast extract (TSB-YE) (Pronadisa, Spain) at 37°C for 20h. Cultures were centrifuged at 2,862 g for 10 min and resuspended in 0.1% peptone. The optical density of each suspension was adjusted to 0.5 (108 CFU/ml) at 600 nm using a spectrophotometer (Spectro UV-Vis; Model UVD-2960, Labomed, Inc., CA). Suspensions were divided into 1ml aliquots, and were subjected in triplicate, on ice to different gamma irradiation doses using 60Co Gamma-irradiator (point source, AECL, IR-79; Technopole, Sidi Thabet, Tunisia). The applied doses were 0.5, 0.7 and 1 kGy. The irradiated cultures were diluted 50-fold using 50 ml of fresh TSB-YE medium and then harvested after 24h of incubation at 37°C with shaking.
2. Determination of morphological changes by AFM
In order to visualize the effect of Gamma-rays on cytoplasmic membrane, L. monocytogenes cells were examined, in triplicate, by atomic force microscopy (AFM) (Nanoscope IIIA, Digital Instrument, VEECO) after 0.5, 0.7 and 1kGy gamma irradiation doses. For this experiment, the cells were collected, washed three times with PBS, and centrifuged. The final pellet was resuspended in PBS, placed on a round microscope cover slide and was simply dried in air according to the method previously described by Braga and Ricci [18].
3. Analysis of fatty acids
To analyze the total cellular fatty acids, Listeria strains were incubated after irradiation treatment in buffered peptone water (Biolife Italiana) for 24 h at 37 °C. Each experiment was performed in triplicate by using three independently grown cultures. Cells recovered from 10 ml suspension were pretreated following the Microbial Identification Inc (MIDI) protocols [19]. All reagents for saponification, methylation, extraction, and washing were dispensed with autopipets into the same tube, making the hands-on time minimal. Next, the final extracts were analyzed by gas chromatography (column: 30 m × 0.25 mm HP-Innowax; flame ionization detect temperature at 280°C; carrier gas N2 at 1 ml/min; injector temperature 270°C; oven temperature programmed from 130 to 230°C) using a Hewlett-Packard HP 5890 capillary gas chromatograph linked to an HP Chemstation integrator. The identification of fatty acid methyl esters was performed by external standards (all purchased from Sigma Chemical Co) submitted to the same processes of manipulation as the biological samples analyzed. Total saturated fatty acids (SFA), total unsaturated fatty acids (UFA) were used to determine the differences among membrane fatty acids of L. monocytogenes cells, and they were examined under the different conditions. The UFA/SFA ratio was used as an indirect indicator of the membrane fluidity.
4. DNA extraction and polymerase chain reaction
Genomic DNA of treated and untreated L. monocytogenes cells was extracted using the bacteria DNA preparation kit (JenaBioscience) as manufacturer’s instructions. The concentration and the quality of the DNA were determined by measuring the OD260nm and the agarose gel electrophoresis (0.8%) containing ethidium bromide. Primers used in both PCR and qRT-PCR are listed in Table 1.

These primers were used in our last studies and by Miladi et al. [13, 39]. Polymerase chain reaction (PCR) was performed in a reaction volume of 25 μl containing 2.5 µl 10X DreamTaq Green Buffer (Thermo Scientific), 100 µM dNTP Mix (JenaBioscience), 0.2 µM of both forward and reverse primers, 1 unit of DreamTaq DNA Polymerase (Thermo Scientific), and approximately 50 ng of DNA template.
PCR reactions were conducted in a MultiGene gradient thermal cycler (Labnet International) using the following program: 95°C for 5 minutes, 35 cycles of 95°C for 30 seconds, 57°C (annealing) for 30 seconds, 72°C for 30 seconds, and finally 72°C for 10 minutes. PCR products were analyzed by electrophoresis on 1.5% agarose gel ethidium bromide stained.
5. Isolation of total RNA
Total RNA was isolated from Gamma irradiated and control L. monocytogenes cells using the SV total RNA isolation system, following the manufacturer’s instructions (Promega, Madison, WI, USA). The RNA was treated with one unit DNase I enzyme (Thermo Scientific) to eliminate residual genomic DNA. Samples were electrophoresed in a formaldehyde agarose gel to check the integrity of RNA.
6. First strand cDNA synthesis
For the reverse transcription (RT) reactions, a mixture of 500 ng of total RNA, 1 µl of 10 µM reverse gene-specific primer, 0.6µl of 100 mM dNTP Mix (Jena Bioscience), 4 μl of MMLV Reverse Transcriptase 5X reaction buffer (Promega), and 200 units of Moloney Murine Leukemia Virus (MMLV) Reverse Transcriptase (Promega) and the final volume was adjusted to 20 μl with Nuclease-free water. The synthesis reaction was carried out for 45 min at 40°C and stopped by incubation at 75°C for 5 min. To avoid detecting genomic DNA contamination in RT-PCR, a control RNA sample without MMLV reverse was used.
7. Real-time PCR assays for relative quantification of gene expression
Real-time PCR was performed using a Chromo4 real-time PCR detection system with Opticon Monitor 3 software (Bio-Rad, Hercules, USA). Amplification and detection were carried out in 96-wells plate with qPCR GreenMaster containing EvaGreen as the fluorescent DNA stain (JenaBioscience). The primers used to perform the quantitative PCR were the same as those used previously for PCR optimization (Table 1). Each reaction was done in triplicate in 20 μl of final volume: 10µl qPCR Green Master, 1 µl of each forward and reverse primers (10 µM), 4µl reverse transcription product as a template and 4 µl PCR-grade water (Jena Bioscience). Amplifications were carried out as follows: 95°C for 2 minutes, followed by 40 cycles of 95°C for 15 seconds, 57°C for 20 seconds and 72°C for 30 seconds. To assess the PCR specificity, the melting curve analysis was performed by increasing the temperature from 50 to 95°C, with a raise in temperature by 1°C every 10 sec with a plate scan step to read the fluorescent signal. The method of Pfaffl et al., 2002 was used to analyze the relative gene expression from real-time quantitative PCR experiments [20].
8. Statistical analysis
Statistical analysis was performed using the S.P.S.S. 13.0 statistics package for Windows. The differences in UFA/SFA ratio were examined by the Friedman test, followed by the Wilcoxon signed ranks test. P-values < 0>
1. Morphological alterations of irradiated Listeria monocytogenes
Cells were scanned after Gamma irradiation treatment (Figure 1). Like the control L. monocytogenes, the 0.5 kGy irradiated Listeria strains have a normal rod shape with a smooth surface (Figure1 A-B); whereas the treated cells with 0.7 kGy gamma irradiation dose present irregular shape and rough surface characterized by dispersed lumps (Figure1 C-D). However, after 1kGy gamma treatment the membrane was destroyed and cell components were scattered around (Figure1 E-F).

2. Membrane fatty acids composition of irradiated Listeria monocytogenes
The membrane fatty acid composition of L. monocytogenes cultured under the different growth conditions (irradiated and normal cells) was determined using a chromatographic method (figure 2).
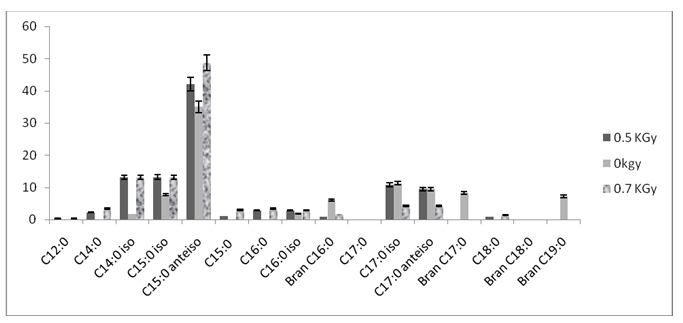
The X-axis represents the 16 detected fatty acids in the studied strain and the Y-axis represents the relative fatty acid abundance (Percentage of total fatty acids).
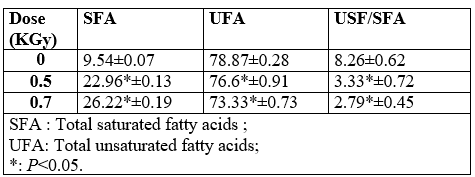
Sixteen fatty acids were found in control and stressed cells. The five main peaks were identified as Tetradecanoic acid (iso-C14:0), Pentadecanoic acid (iso-C15:0 and anteiso-C15:0) and Heptadecanoic acid (iso-C17:0 and anteiso-C17:0). Their relative percentages were between 1.68% for iso-C14:0 and 35.02 % for anteiso-C15:0. As expected, the irradiation of L. monocytogenes resulted in differences in membrane fatty acid composition. Indeed, a significant increase in SFA accompanied by a significantly decrease in the UFA was observed (P < 0>
It has been previously reported that membranes with high UFA/SFA ratio show a high fluidity [21, 22].
3. Virulence genes expression
Relative [removed]RE) of prfA, fri and hlyA genes were determined after exposition of Listeria monocytogenes cells to low-dose gamma irradiation (0.5 kGy and 0.7 kGy). To measure transcripts levels of these virulence genes, various steps in the qRT-PCR procedure were optimized. The RNA sample quality remained stable over the period of analysis and showed no differential transcript degradation. Primers for individual genes were chosen to be specific for L. monocytogenes. In addition, the melting curve was carried out to detect any no specific amplification.
The optimization of the real-time PCR was evaluated based on the amount and the quality of total RNA template, specificity and efficiency of the reaction. The observed results suggest that the conditions used for RNA template preparation, reverse transcription and real-time PCR were compatible with quantitative analysis of RNA transcripts.
Expression levels were evaluated after the comparison of the two conditions (control and gamma treated cells).
The analysis of virulence gene expression in L. monocytognes using Real-Time PCR revealed that both genes prfA and fri were upregulated after low doses Gamma exposition. While hlyA gene was mostly down-regulated (Figure 3).

In addition, the expression levels were more pronounced after gamma treatment with 0.7 kGy dose. Indeed, the fold changes expression of prfA in treated L. monocytognes cells were 3.22 and 59.79 after 0.5 kGy and 0.7 kGy gamma doses, respectively. Similarly, fri gene was overexpressed after 0.5 kGy and 0.7 kGy in L. monocytognes (2.07 and 10.06 fold change, respectively). On the other hand, the hlyA gene was down-regulated in treated L. monocytognes cells.
Thus, the relative expression of prfA, which is a regulator gene may depend on gamma radiation dose and influences the expression of the two other studied genes.
In this study, three low radiation doses were chosen which are generally tolerated in food irradiation to eliminate pathogenic bacteria. Various factors, such as chemical composition of foods may affect the radiation resistance and sensitivity [5, 7]. Thus, as a try to reduce this problem, gamma treatment was applied for bacteria incubated in TSB-YE.
The effectiveness of gamma irradiation in inactivating pathogenic bacteria at low gamma doses was previously reported; E. coli 0157:H7 and aerobic mesophiles in lettuce at 0-0.55 kGy [23], L. monocytogenes and S. enterica growth was inactivated in fresh orange juice at 0-4 kGy [15], S. typhimurium and E. coli in carrot and kale juice at 0-3 kGy [7] and total aerobic and coliform bacteria in carrot and kale juice were inactivated at 0-5 kGy [5].
Some radiation-resistant bacteria can recuperate their growth potency when the environmental conditions are suitable for bacteria development. In our study L. monocytogenes strain was resistant to 0.5 and 0.7 kGy gamma irradiation dose but not to 1kGy.
The 0.5 kGy dose did not significantly affect neither the shape nor the smooth of the bacteria.
Bacterial cytoplasmic membranes define cells from the external environment. It represents an additional location for stress sensing that is capable to transduce signals affecting the expression of genes involved in defense [24]. It has been proposed that under some stress conditions like starvation, member of L. monocytogenes can undergo a number of morphological and physiological changes to adapt to such severe condition [25]. According to the obtained results, important modifications seemed to take place after gamma irradiation treatment with 0.5, 0.7 and 1 kGy dose for L. monocytogenes. This sensitivity implies that membrane lipids play an important role in microbial adaptation to the destructive effect of gamma rays. We also observed that the bacterial membrane resistance was associated with the alteration of virulence gene expression. According to Russel [26], a change in membrane fluidity is often caused by a change in the lipid composition of the membranes to keep an adequate proportion of liquid-crystalline lipid in the membrane and to maintain the bilayer phase. In the same way, treatment of E. coli with non-lethal doses of heat or benzyl alcohol causes transient membrane fluidization and permeability, and induces the rapid transcription of heat-shock genes in a σ32-dependent manner [27].
Total saturated and unsaturated fatty acids were used to determine the differences among membrane fatty acids of L. monocytogenes cells grown under the different stress conditions. The UFA/SFA ratio was used as an indirect indicator of the membrane fluidity. Significant modifications on fatty acid (FA) composition (P ≤ 0.05) seemed to take place after Gamma-irradiation treatment with a 0.5 kGy and 0.7 kGy doses for studied strain. So, a significant (P ≤ 0.05) decrease in the UFA/SFA ratio was observed with the two doses. Moreover, our results showed a novo synthesis of membrane lipids: C12:0; C14:0; C15:0; C16:0 and C18:0. A wide range of bacteria are able to change their membrane composition by novo synthesis of membrane lipids during growth [28]. Bacteria regulate their lipid composition to achieve a degree of fluidity compatible for life. However, little is known on the influence of these modifications in membrane composition on the bacterial resistance to subsequent stresses. One of the most important consequences of membrane fatty acid changes in microorganisms is to modulate the activity of intrinsic proteins that perform functions such as ion pumping and nutrient uptake [29]. The membrane lipid plays an important role in microbial adaptation to the destructive effect of the reactive oxygen species. This adaptation was related to the increase of SFA production. Therefore, it’s supposed that the modification of SFA proportions constituted an adaptive response to Gamma-irradiation stress in L. monocytogenes. It is important to note that several studies have suggested that an increase in SFA content could cause a decrease in membrane fluidity [21, 22].
Listeria monocytogenes responds to environmental stress by modifying the rate of synthesis of certain proteins [30]. This study showed modification on relative expression of Listeria monocytogenes genes after its treatments with Gamma rays. The mechanism of microbial inactivation by gamma ray may damage DNA and/or some protein denaturation [31]. For a living cell, the slightest damage to DNA can affect its development and survival. Thus, gamma lesions impinge on vital cellular functions, including transcription, DNA replication and cell cycle progression. Their persistence in the genetic material also increases the chance of fixation into mutations [32]. These damages may affect directly microbial DNA by making strand breaks of nucleic acids or indirectly via hydroxyl radicals originating from radiolysis of water [33]. In our study, the expression of pathogenic genes was affected after 0.5 and 0.7 kGy gamma irradiation (figure 3). The qRT-PCR analysis demonstrated that prfA and fri genes were up-regulated after 0.5 kGy treatment and were strongly induced after 0.7 kGy dose. However, the hlyA gene was down regulated. Other genes could be affected by this treatment and their expression could be influenced by the product of prfA expression. It has been previously demonstrated that the expression of 145 L. monocytogenes genes is influenced by PrfA according to transcriptomic profiling and proteomic analyses [34-36]. These indirectly regulated genes include transporters, metabolic enzymes, regulators, proteins of different functions, together with many members of stress response regulation [34, 35]. Indeed, PrfA is a central virulence regulator and may potentially exert many general effects on L. monocytogenes homeostasis. Although loss of expression from the P2prfA promoter affects in vitro virulence phenotypes, such as hemolysin production [37].
It has been reported that gamma irradiation could induce gene mutation. But, there has been no evidence of irradiation-induced enhanced pathogenicity of food borne microorganisms. However, it should not be ignored that there were no involved mechanisms of post-irradiation DNA repair.
Lim et al. [38] reported that after exposure to a dose of 0.5 or 1 kGy, the transcriptional changes of vvhA, ctxA, and tdh genes respectively in V. vulnificus, V. cholerae, and V. parahaemolyticus ranged from 0.3- to 0.001-fold. He also indicated in his study that gamma radiation reduces the virulence gene expression of surviving pathogens.
Several genes contributing to L. monocytogenes virulence have been well identified and characterized. The majority of these virulence genes, lies in a single cluster on the L. monocytogenes chromosome, are controlled by the transcriptional activator PrfA [39, 40]. This is the first report mentioning the effect of gamma irradiation on L. monocytogenes gene expression.
Beauchamp [11] has compared the natural resistance of the genome of Gram-positive and Gram-negative bacterium against irradiation solution of pure DNA and bacterial strains such as Listeria monocytogenes and Escherichia coli which were irradiated using gamma rays. The authors demonstrated that gamma rays induce the formation of cyclobutane pyrimidine dimers and 6-4 photoproducts in DNA in L. monocytogenes genome between 0.1 and 0.7 kGy gamma irradiation doses. In the other hand, it’s supposed that the combined effects of both stresses (freezing and ionization) disrupted the expression of the studied genes. In previous study, it was demonstrated that simultaneous exposure to most multiple stresses (e.g., osmolarity, acidity and temperature) resulted in higher resistance of L. monocytogenes at pH 3.5 than exposure to the same stresses sequentially [41]. Similar results were found with a combination of acid , water activity (aw) and temperature stress [42]. These results strongly support our expectations.
The main aim of this paper was to demonstrate whether the gamma rays can affect Listeria monocytogenes cells. The first observation was that this treatment leads to in typical form modification of Listeria. Then, by measuring the relative expression of three virulence genes it was demonstrated that low gamma doses are able to induce pathogenic gene expressions. Moreover, prfA expression was very sensitive to gamma rays and its induction was strongly related to other virulence genes expression.
Further studies are needed to explain mechanisms for regulating expression of irradiation-stress response and determining the effect of such treatment on food products in order to limit their application in food industry.
This research was supported by the National Center for Nuclear Sciences and Technologies (CNSTN), Tunis Cedex, 2020, of Tunisia. The authors thank Pr. Mokhtar Hamdi for the irradiation essays.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
