
Review Article | DOI: https://doi.org/10.31579/2637-8914/227
1Department of Food Science, College of Agriculture and Food Sciences, Florida A and M University, Tallahassee, FL 32307, USA
2Whistler Center for Carbohydrate Research and Department of Food Science, Purdue University, West Lafayette, IN 47907, USA
3Department of Nutrition and Integrative Physiology, Florida State University, Tallahassee, FL ,47907, USA
*Corresponding Author: Oluwatoyin O. Sangokunle, Department of Food Science, College of Agriculture and Food Sciences, Florida A and M University 1740 Martin Luther King Jr., Blvd, FL 32307, USA.
Citation: Oluwatoyin O. Sangokunle, Anna M. R. Hayes, Sarah G. Corwin, (2024), Assessing slowly and resistant starches from pulses for nutrition and health, J. Nutrition and Food Processing, 7(6); DOI:10.31579/2637-8914/227
Copyright: © 2024, Oluwatoyin O. Sangokunle. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 08 April 2024 | Accepted: 20 May 2024 | Published: 06 June 2024
Keywords: pulses; microbiota; prebiotics; starch; resistant starch; fermentation
The pulse protein market is growing at a 6.0% annual compound growth rate (2021 – 2028) due to the popularity of vegetarianism, leading to an increased opportunity to utilize its starch. Pulse starch contains significant amounts of slowly digestible starch and resistant starch, both of which have been characterized and have the potential to contribute positively to human health. However, applications of pulse starch related to digestibility and fermentability patterns and physiological effects on human health remain largely unexplored. This review aims to provide background findings, summarize data on pulse starches concerning their digestibility and colonic fermentability, and discuss the potential application of pulse starch benefits to human health.
In the quest for plant-based proteins suitable for the growing meat analog market, pea protein is a key player, and other pulse proteins are showing considerable growth with a compound annual growth rate of 3.15% for chickpeas, 4.0% for fava beans, and 3.2% for chickpea in 2028 (Mordor Intelligence, 2021; The Sentinel, 2021). Commercial pulse protein production generates 35-60% starch as a side stream byproduct. With the rising demand for pulse protein, there will be a considerable increase in unutilized starch. In the call for a more eco-friendly society and the increased demand for plant-based products, there is a need to reduce the food waste generated from pulse protein production and improve its utilization as a healthy, functional ingredient.
A step towards pulse starch utilization and its transformation into novel functional ingredients for food and human health is a promising new avenue. Nutritionally, some isolated pulse starches are an excellent source of slowly digestible starch (SDS) and can potentially be a source of resistant starch (RS) (Lu et al., 2018). Moreover, pulse starch has superior physical functionality, as it has been reported to have good heat and mechanical shear stability and can be used in high-temperature and high-mechanical food systems (Singh, 2011). SDS is beneficial to human health as it has been shown to decrease glycemic response, improve hunger and satiety responses, and has the potential to prevent and ameliorate obesity (Lim et al., 2021). RS is the undigested portion of starch that reaches the large intestine, is primarily fermented, and is used as an energy substrate by gut microbes, producing short-chain fatty acid (SCFA) metabolites that impart health benefits to the host. Studies have demonstrated multiple health benefits associated with increased consumption of foods formulated with SDS and RS (Ble-Castillo et al., 2017; Bodinham et al., 2014; Meenu & Xu, 2019). Consumption of RS has been shown to improve bowel movement, weight management, insulin regulation, and prevention and control of colon cancer and obesity (Meenu & Xu, 2019; Peterson et al., 2018). Further, RS promotes and causes diet-induced changes to the types of bacterial communities in the gut (Birt et al., 2013; Higgins & Brown, 2013; Keenan et al., 2015). The beneficial effects of RS have prompted rising interest in its use in foods related to nutrition and health. Accordingly, the use of RS as a food ingredient is becoming more prominent, with a projected growth rate of 4.9% from 2020 to 2025 (Market Growth Report, 2020).
In this review, we focus on isolated, purified starch as recovered as a byproduct from the production of pulse protein. This review highlights the potential nutritional and health prospects for expanded utilization. The aim is to describe the possible use of pulse starch, a major waste stream of pulse protein production as a slowly digestible ingredient, the physiological effects of pulse starch on human health, and how pulse RS can potentially be used to promote healthy gut microbiota.
Starch is a renewable polymer formed from glucose and is a sustainable carbon source for plant growth (Sulpice et al., 2009). Starch consists of linear and branched glucose polymers known as amylose and amylopectin, respectively. Glycosidic bonds link the glucose polymer chains with α-glucose monomers bonding at carbon positions 1 and 4 (α-1,4 linkages), constituting linear polymer chains and those bound through carbon at positions 1 and 6 (α-1, 6 linkages), creating branch points that are further elongated by α-1,4-linked linear chains. The type and placement of these linkages affect starch digestibility and function.
Starch structures in their native crystalline states exist helically (F. Zhu, 2018). Both amylose and amylopectin can exhibit single and double helical configurations. The hydroxyl group and hydrogen bonds of glucose residues in the helical configuration are outwardly and inwardly positioned, respectively. This positioning makes the hydrophilic and hydrophobic regions well-defined, which in turn dictates the surface interactions of native starch. Native starch is generally insoluble in water but can interact with other substances and elements such as lipids and iodine. The lipid-starch complexes are naturally present in some native starches and can be found externally at the surface of the granule or internally (inside the starch granule) (Putseys et al., 2010). The iodine-amylose complex is based on the ability of iodine to penetrate the core of the left-handed helix, which results in visible color development during the starch-iodine test (blue-black coloration). This complex forms a single left-handed helical structure of amylose (Putseys et al., 2010).
Amylose is a long linear chain of glucosyl residues linked by α-1,4 bonds, while amylopectin is a shorter linear-branched chain of glucosyl residues branched by α-1,6 bonds (Seung, 2020) (Figure. 1).

Figure 1: Illustration of starch digestion
The branched amylopectin is arranged in a clustered structure of adjacent chains resulting in double helices. The helices form the crystalline region of the starch granules, visible under light microscopy. X-ray diffractometry is used to show the degree of crystallinity of starch granules. The chain length of amylopectin determines the starch granules’ crystalline or polymorphous state. Starch exists in two major semi-crystalline polymorphous structures in its native state, which were observed using X-ray diffraction, A and B polymorphs. In addition, a third identified intermediary type polymorph, type C (Pozo et al., 2018). Starch type C is a combination of types A and B. The type A, B, and C classifications under X-ray diffraction correlate with the amount of amylose and amylopectin. Type A and B starches are usually observed in cereals, roots, and tubers. In contrast, type C is most common in unripe bananas and pulses (dry beans, peas, and lentils) (Ashogbon et al., 2021). The ratio of distinct amylose and amylopectin, combination and distribution, and the presence or absence of either amylose or amylopectin (in cases of modified starch) are the determining factors for the physicochemical function and nutritional prospects for the starch (Seung et al., 2015).
Starch undergoes paste formation when heated in excess water at a specific temperature through gelatinization and pasting. During cooling, molecular interaction occurs first between amylose polymers, then later between amylopectin polymers, and between amylose and amylopectin, forming ordered structures (Chang et al., 2021). The process of forming a reordered structure is termed retrogradation. Retrogradation is associated with gel firmness, increased crystallinity and is linked to the rate of starch digestibility.
2.1 An overview of starch digestion and nutritional classifications
The starch digestion process starts with a mixture of salivary α-amylase in the mouth, followed by the action of intestinal pancreatic α-amylase in the small intestine. The resulting oligomers (mainly maltose and maltotriose) and α-limit dextrins (low molecular weight branched hydrolyzed starch products) of the digested starch are further hydrolyzed to glucose by the α-glucosidases at the brush border of the small intestine (Brownlee et al., 2018) (Fig. 1).
Depending on the rate of starch digestion and conversion to glucose, its absorption results in low or high glycemic and insulinemic responses. Understanding how starches differ in glycemic and insulinemic responses is essential for developing pulse starches with enhanced, slowly digestible, or resistant properties (Zhang & Hamaker, 2009).
The nutritional classification of starch is based on the rate of in vitro digestion and has been classified by Englyst et al. (1992) into rapidly digestible starch (RDS), SDS, and RS. The Englyst in vitro method has limitations and is designed to mimic human digestion of starch using pancreatic α-amylases and amyloglucosidase. It is based on the rate of hydrolysis of starch to glucose with continuous agitation at 37°C. An aliquot of the digested starch mixture is sampled at the starting point, followed by glucose quantification at 20, 120, and 240 min, where glucose is measured using the glucose oxidase-peroxidase method. RDS is starch digested within the first 20 min, SDS between 20 and 120 min, and RS is the amount of starch undigested at 120 min. Starch structure interferes with the digestive enzymes and is the basis for RS nutritional classifications. It is primarily determined by in vitro methods compared to in vivo due to variations and complexity associated with the latter. In-vivo methods of starch classification typically involve analyzing the behavior and effects of starch in living organisms, often through controlled experiments or observational studies. One common approach is to examine the glycemic response elicited by different starches when consumed by human subjects. The glycemic response refers to the increase in blood glucose levels following the consumption of carbohydrates, including starches. This response can vary depending on the type of starch, with some starches causing a rapid increase in blood glucose levels (high glycemic index) and others leading to a slower, more sustained increase (low glycemic index).
One in-vivo method used to classify starches is the determination of glycemic index (GI), which measures how quickly a carbohydrate-containing food raises blood glucose levels compared to a reference food, typically glucose or white bread (Atkinson et al., 2008, 2021). Starches with a high GI are rapidly digested and absorbed, leading to a quick spike in blood glucose levels, while those with a low GI are digested and absorbed more slowly, resulting in a more gradual increase in blood glucose levels. Although the in vivo method is robust and gives a better RS prediction than the Englyst in vitro, it is time-consuming and expensive (Martens et al., 2019; Regina et al., 2006; L. J. Zhu et al., 2011).
SDS has been associated with health benefits such as lowering glycemic index or response, whereas RS has been shown to promote healthy gut bacteria growth (Voreades, Kozil, & Weir, 2014). Both SDS and RS can provide a dietary means of reducing the risk of cardiovascular, metabolic, and gastrointestinal diseases (Clemente & Olias, 2017). Previous studies have used processing methods to increase the SDS and RS contents of pulse starches to facilitate the application of pulse SDS and RS. Chung, Liu, and Hoover (2010) reported increasing levels of SDS and RS in lentil, navy bean, and pea starches after annealing and moist heat treatments. Similarly, maltogenic alpha-amylase treatment on pea, faba bean, and lentil starches increased RS levels by 4, 6, and 6%, respectively (J. Li et al., 2021). Some studies have shown complete fermentation with high SCFA production after 24 h of in vitro fermentation (Brummer et al., 2015; Guan et al., 2020; P. Li et al., 2019; Xiong et al., 2018). We have also found that some cooked pulse starches exhibit slow digestibility properties, and their respective RSs are completely fermentable with high acetate and butyrate production and promote beneficial bacteria (O. O. Sangokunle & Sathe, 2024). Moreover, beneficial butyrate-producing bacteria were promoted during this fermentation of RS at 24 h. Hence, developing SDS and RS from pulse starches to promote healthy glucose homeostasis and the growth of healthy gut bacteria can be a good use of the pulse protein waste stream.
Pulse starch granules have no surface pores (Fig. 2), contributing to slow digestibility in their raw state (Fig. 2). Absence of surface pores on pulse starch granules decreases enzymatic access during hydrolysis of raw pulse starch (Chung & Liu, 2012; Hoover et al., 2010; Simsek et al., 2016). In our previous findings, cooking purified pulse starch increased its digestibility, causing a rapidly increased level of RDS and reduced levels of SDS and RS. Rapid retrogradation occurs during the cooling (24-48 h, 4°C) of pulse starches, causing increased resistance to digestive enzymes, as indicated in bread (unpublished data).

Figure 2: Scanning electron micrographs of chickpea starch granules showing no surface pores at 3,000 and 10,000 x magnifications
3. 1 Short description of how to make SDS
It is well known that most gelatinized waxy starches, processed starchy foods, and foods high in α-1,4 linkages. Raw starch granules, retrograded amylose, and some physically or chemically modified starches are resistant to digestion due to physical or chemical characteristics. Purified pulse starch can be made slowly digestible through decreasing accessibility of enzymes to the starch, chemical structure limiting the enzymatic rate of action, and non-starch enzyme inhibitors (Lee et al., 2013; Zhang & Hamaker, 2009). In contrast, α-1,6 glycosidic bonds are typically more slowly cleaved by intestinal brush border glucosidic enzymes, and in general, an increase in α-1,6 linkages results in a slower rate of digestion (Ao et al., 2007). In gelatinized native starches, the ratio of α-1,4 to α-1,6 linkages is crucial for determining the digestion rate. In vitro experiments have shown that the disaccharides with α-1,3, α-1,2, and α-1,1 also have lower rates of digestion than α-1,4 but faster than α-1,6 (Lee et al., 2016). Further understanding is needed to utilize this information in oligo- and polysaccharides such as purified pulse starch (Lee et al., 2016).
Starch chain length and degree of branching contribute to changes in digestion rate, with increased branching and shorter outer linear chains resulting in slower digestion rates (Ao et al., 2007). Ambigaipalan et al., (2014) reported no changes in chain length distribution (CLD) of amylopectin of black bean, faba bean, and pinto starches after heat-moisture treatment (HMT), due to the structural reorganization and crystalline stability of purified pulse starch, therefore, capable of preserving its slow digestion property. Other studies have also shown an increase in the SDS and RS after HMT of pulse flour (Chávez-Murillo et al., 2018). Physical structures that contribute to slow digestion rates include increased crystallinity through retrograded structures and protein or alginate networks that limit the accessibility of enzymes to bonds between glucopyranosyl units. Apart from structural reorganization and crystalline stability of purified pulse starch structures, inhibitors such as polyphenols have previously been shown to generate a slow digestion property in maltodextrin, waxy corn, and corn starch through their action on enzymes, as opposed to a change in the structure of the carbohydrate (Lim et al., 2019; Yu et al., 2021). This approach can also be used to generate a slow digestion property of purified pulse starches through their action on amylolytic enzymes, as opposed to a change in the structure of the starch.
3.2 Resistant starch
Resistant starch (RS) is often found in starchy food such as cereals, pulses, roots, and tubers. RS is present or included in finished food products such as noodles, pasta, white bread, breakfast cereal, cakes, tapioca and yam flours, cooked potatoes, baked bread or cake, and cooked rice (Bello-Perez et al., 2020). The RS content in purified raw pulse starch decreased after cooking (L. Li et al., 2019). As reviewed elsewhere, in whole pulse seeds, starch incorporation into the seed matrix limits digestive enzymes from penetrating the starch surface, but this phenomenon does not occur in purified starch (Ajala et al., 2023). The starch fraction should be separated from non-starch components to produce quality pulse RS from the co-produced starch during pulse protein production.
Starch is co-fractionated during pulse protein production, followed by air classification (dry milling) or sedimentation and centrifugation (wet milling). The separation process of the starch fraction from pulses has been reported by various researchers (Jiang et al., 2019; G. et al., 2018; Sangokunle et al., 2020). The wet milling method is commonly preferred due to the high purity of the final product (Sangokunle et al., 2020). The goal of starch isolation from seed matrix and flour is to ensure the complete removal of other non-starchy components by repeated washing or classification depending on the isolation method. Isolated starch is often pre- or post-treated with alcohol, alkaline, or a combination of both to remove non-starchy components such as protein and surface lipids (O. O. Sangokunle et al., 2020). We have observed mucilaginous material and residual fiber co-sedimented with isolated pulse starches using wet milling, posing questions on pulse starch purity. The effects of these starch purification treatments on pulse RS and their influence on starch digestibility still need to be investigated.
Generally, resistant starches (RSs) are structurally classified based on physical inaccessibility to digestive enzymes (RS1), nature of the starch granule (RS2), food processing conditions causing retrogradation of amylose and amylopectin (RS3), chemical modification (RS4), and amylose-lipid complexation (RS5). A starch’s structure can interfere with accessibility by native digestive enzymes (salivary α-amylase, pancreatic α-amylase, and the small intestinal α-glucosidases). Various percentages of raw and cooked RS in pulses have been quantified using the in-vitro method, as shown in (Table 1). Raw purified garbanzo bean starch (9%) and navy bean starch (77%) have the highest reported RS, respectively. In an in-vitro method, RS is classified as the starch fraction recovered after 4 h hydrolysis with digestive enzymes (alpha-amylase and amyloglucosidase). Pulse RS can be produced from purified pulse starch by structural modifications of native starch to prevent accessibility to amylolytic enzymes. These modifications can be carried out by physical, chemical, and biological treatment of the starch or a combination of these treatments.
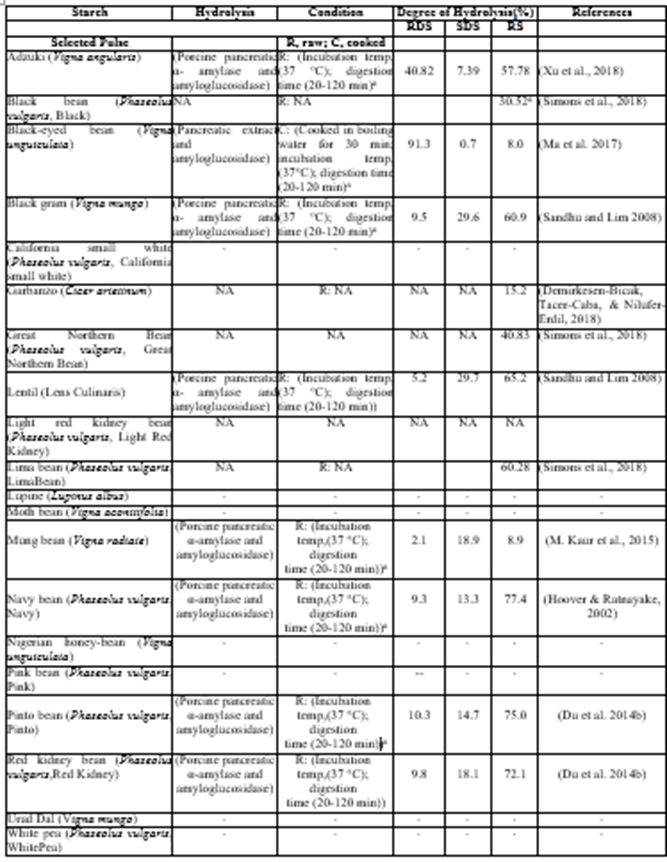
Abbreviations: C, cooked; NA, not applicable; R, raw; RDS, rapidly digestible starch; RS, resistant starch; SDS, slowly digestiblestarch.
Table 1. Compilation of examining digestion characteristics of pulse-based starches.
3.3 Starch modifications to increase RS
Some purified raw pulse starch is generally known to contain RS2 as it is structurally resistant to digestion due to its native state. Additionally, pulse starches contain significant amounts of RS1 in their processed form, as affected by mastication (Alpos et al., 2021). Although RS from purified pulse starch has yet to gain utilization in commercial food applications compared to corn, banana, and potato starches. Some researchers have identified, characterized, and modified novel RS from a whole seed or flour as a potential functional ingredient (Morales-Medina et al., 2014; Ovando-Martínez et al., 2011).
Generally, starch is physically modified by HMT, annealing, or other forms of heat treatment such as irradiation, autoclaving, boiling, parboiling, and microwaving to alter the structural arrangement of amylose and amylopectin for digestibility (Bello-Perez et al., 2020). Physical modification is usually conducted below the starch gelatinization temperature, often coupled with rapid or slow cooling (tempering) to produce a retrograded starch which in turn increases crystallinity and causes a decrease in the rate of digestion (Patel et al., 2017). We pose that these processes, either alone or in combination, could be studied for their ability to produce RS with desirable functional properties in pulse starch.
Chemical modification of starch is usually conducted by the addition of hydrocolloids such as gums or lipids. Starch, mostly amylose, forms inclusion complexes with lipids and surfactants while reordering, resulting in aggregated semi-crystalline starch. This interaction prevents access of amylolytic enzymes. This type of chemical modification has been reviewed previously (Parada & Santos, 2016). For example, an average of 15 percent decrease in digestibility has been reported in potato starch hydrolysis after cooking in 0.5 percent guar gum (Bordoloi et al., 2012). Similarly, during the first (10 min) and at the end (240 min) of in vitro digestion of waxy-maize starch-guar gum complexes, respectively, Dartois et al. (2010) reported (25 percent) and (15 percent) decreases in digestion. These modification methods can be utilized for purified pulse starches to enhance their utilization.
Starch modification by enzyme involves using α-amylases and debranching enzymes, such as pullulanase or isoamylase, and recrystallization. Pullulanase or isoamylase is used to generate small linear chains from the branched amylopectin of the starch (Liu et al., 2017). The obtained mixture of debranched starch has been reported to indicate a decrease in amylolytic hydrolysis and an increase in RS content (Liu et al., 2017). Increasing amounts of RS have been observed in gelatinized waxy maize starch subjected to a high pullulanase treatment (20-40 ASPU/g) and then recrystallized (Miao et al., 2009). In general, starch granules, starch crystallinity, amylose chain length, the ratio of amylose to amylopectin, amylose retrogradation, and interaction of starch with other biomolecules can influence the production and formation of RS. Enzymatic starch modification methods could increase RS in pulse starches, thus improving physiological benefits from their consumption.
3.4 Commercial adaptability of resistant starch from pulses
Commercially, RS from pulse starches is not commercially available. In contrast, RS from corn, including Hylon VII, Amylomaize VII, and Novelose 240, is produced in the United States by the Ingredion and National Starch and Chemical Company (Haralampu, 2000). The food industry utilizes modified starches for the functional properties they provide to finished food products. RS performs better than traditional fiber by providing distinct sensory product qualities and desired organoleptic properties with no impact on taste. RS generally offers excellent gel formation and high water-binding capacity with increased swelling (Baixauli et al., 2008). Recently, due to the increasing worldwide need for food and the trend towards “clean label” food products, alternative options to meet the demand for natural and sustainable food ingredients such as pulse starch are critical. Also, the use of starch generated from commercial pulse protein production is crucial to mitigate food waste and increase sustainability. Several companies sell pulse-based proteins as a form of sustainable, clean-label protein; however, to our knowledge, no food company currently offers pulse-based RS products commercially. It is possible that the starch extracted when purifying the protein from pulses could also be used, thereby eliminating a waste stream. Some of this starch could be modified into RS, or it may naturally be RS, constituting an additional product with added value.
3.5 Fermentation of pulse RS and gut metabolites
Incorporating pulse RS in the diet can modulate gut microbiota and promote health. Pulse flour has been shown to directly promote butyrate and acetate production (supporting immune health and homeostasis) and encourage the growth of genera Bifidobacterium, Clostridium, Eubacterium, Faecalibacterium, and Roseburia (Gullón et al., 2015). In vitro, fecal fermentation studies of cooked and raw bean polysaccharide extracts isolated from four Phaseolus vulgaris L. cultivars have shown acetate to be the most abundantly produced SCFA for all cultivars (Ze etal., 2012). Our recent investigation found that acetate and butyrate are the most produced SCFAs during 24 h fecal fermentation of purified pulse starches (O. O. Sangokunle & Sathe, 2024). There is a need for further assessment of health benefits associated with RS of pulse origin,mainly from purified pulse starches.
Prebiotics are defined as “a selectively fermented ingredient that results in specific changes in the composition and activity of the gastrointestinal microbiota, thus conferring benefit(s) upon host health” (Roberfroid et al., 2010). Evidence suggests that RS supplementation as prebiotics (but not specifically from pulse starch) has been shown to improve postprandial glycemic and insulinemic responses, reduce cholesterol and triglycerides, increase satiety, reduce fat storage, and promote weight loss, making it an exciting ingredient for the management of metabolic disorders (Ai et al., 2013; Higgins, 2014; Wang et al., 2019). Notably, most studies use RS from cereal or tubers sources, and there is a lack of data regarding the prebiotic benefits of RS isolated from pulse starches. Interestingly, Yadav et al. (2010) compared the RS content of cereals, legumes, and tubers, which were conventionally boiled and pressure-cooked. Data from the study indicated that legumes generally had higher RS content than cereal and tubers, with lentils having the highest RS content amongst all tested foods. Siva et al. (2019) evaluated differences in dietary fiber in pulses and found 12%, 15%, and 12% dietary fiber in lentils, common beans, and chickpeas, respectively. Although RS likely makes up for a good portion of the dietary fiber found in these pulses, the proportion of distinct dietary fiber components to total RS content in each source was not estimated. RS content in foods is not only determined by intrinsic starch properties (e.g., crystallinity, amylose to amylopectin ratio, granular structure). However, processing procedures also strongly influence it (Ai et al., 2013). In brown lentils (Lens culinaris, Medikus), RS content was shown to be enhanced by the addition of lipids, which results in the formation of amylose-lipid complexes that are resistant to digestion (RS Type V) (Okumus et al., 2018). On the other hand, other processing techniques that can disrupt the native starch macrostructure contribute towards increased accessibility of these starches by digestive enzymes in the upper gastrointestinal tract. Thus, this could reduce the RS content in pulses, as previously reported (Kasote et al., 2014).
Finally, a few cited literatures compare RS content in pulses and how it is affected by processing techniques. However, there is a lack of studies that explore in detail the physicochemical characteristics of isolated pulse starches from other distinct botanical sources (in their native and processed forms) and how they modulate microbial shifts in the large intestine. Since small shifts in the physicochemical structure of RS can shift gut microbiota composition and metabolism (Deehan et al., 2020), an investigation of the prebiotic effects of naturally occurring and processed RS of pulse origin is needed to characterize their full prebiotic potential.
Studies investigating the physiological impacts and health outcomes of pulse-based SDS and RS still need to be investigated. Given the potential beneficial effects of SDS and RS described above, this is a promising area for future research. There is a need for additional studies on pulse SDS and RS production, susceptibility of pulse RS to amylolytic hydrolysis, effects of pulse RS on colonic health, and glycemic index. Furthermore, improved functional properties of different pulse RS that correlate to their structural properties require elucidation.
CLD, chain length distribution; HMT, heat moisture treatment; RDS, rapidly digestible starch; RS, resistant starch; SDS, slowly digestible starch; SCFAs, short-chain fatty acids
Oluwatoyin Sangokunle: Conceptualization, Writing – Original draft preparation
Anna M.R. Hayes: Writing – review and editing
Sarah G. Corwin: Writing – review and editing
No data has been reported for this article.
The authors declared they have no conflict of personal or financial interest.
Dr. Oluwatoyin Sangokunle thanked the Whistler Center for Carbohydrate Research and the Department of Food Science, at Purdue University for research training support, and Florida State University and the Department of Nutrition and Integrative Physiology, Florida State University, for providing graduate student assistantship.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
