
Research article | DOI: https://doi.org/10.31579/2637-8914/036
*Corresponding Author: E.R. Amante, Food Science and Technology Department, Federal University of Santa Catarina, Florianópolis, SC, Brazil
Citation: J.M. Pereira, A.C.M de S. Aquino, L.J. Dallagnol, E.R. Amante (2021) A Natural Antifungal from Cassava Starch Fermentation Wastewater. J. Nutrition and Food Processing, 4(1); DOI:10.31579/2637-8914/036
Copyright: © 2021 E.R. Amante, This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 14 January 2021 | Accepted: 15 February 2021 | Published: 26 February 2021
Keywords: wastewater; antifungal activity; botrytis cinerea; monilinia fructicola; colletotrichum gloeosporioides; fungi and cassava starch
Sour cassava starch production process, during the fermentation step, an acidic wastewater is generated, in this study named wastewater of cassava starch fermentation (WCSF). It has been studied the effect of WCSFs (from three industrial source: WCSF 1, 2 and 3) and patterns of organic acids (lactic, acetic, propionic and butyric acids, which were the main organic acids present in the WCSFs) singly or together on the development and growing of Botrytis cinerea, Monilinia fructicola and Colletotrichum gloeosporiodes. WCSFs showed antimicrobial effect on the three fungi studied. WCSF2 inhibited the germination of conidia from the three fungi, while WCSFs 1 and 3 inhibited conidia germination of B. cinerea and C. gloeosporiodes. The acetic, propionic, butyric and the mix acids were also effective in the reduction of conidial germination. In the analysis of mycelia growth, it has been observed that the growing was statically smaller, in relation to negative control, for all fungi in the treatments with the WCSFs and with the pattern solution of butyric acid. Which suggest that the WCSFs compounds and butyric acid are effective inhibitor for fungi and justify future studies using this wastewater.
Post-harvest diseases are among the major causes of fruit loss, and microbial growth on surfaces is the most common cause of deterioration of processed fruits (Torres and Karel, 1985; Artés et al., 2007). The most frequent and important pathogens associated to fruits decay are: Monilinia fructicola, Lasiodiplodia theobromae, Colletotrichum gloeosporioides species complex, Cladosporium spp., Rhizopus sp., Mucor spp., Alternaria sp., Botrytis cinerea, Phoma sp. and Penicillium sp. (Callan & Carris, 2004; Hansmann & Combrink, 2003).
The occurrence of these pathogens contributes to compromise the quality of agricultural products has also led to the consumption of pesticides in order to protect the agricultural production and guarantee supply (Damalas & Eleftherohorinos, 2011). However, protecting the orchard and crops, with reduced use of pesticides is an important challenge in defense of the environment and consumer health.
Research and development of natural products to be used as pesticides to control the deterioration of fruits by fungi in the post-harvest period should be encouraged, since toxicity is proven through their consumption for many years (Damalas & Eleftherohorinos, 2011; Chan, 2013). Due to the safety that natural products can represent for animals and the environment, research and development of a new product can represent a much shorter cost and time for registration and making available on the market, compared to a synthetic cemical product (Dukare et al., 2019).
The production process of sour cassava starch involves the natural fermentation of cassava starch and produces a liquid residue, rich in organic acids, traditionally used in food preservation, the main one being propionic acid, but lactic, acetic and butyric acids they can also have a protective action on food (Galvez, Burgos, López, & Pulido, 2014) However, this wastewater, which has not been studied yet as crop protection agent, is, until now, released directly into the environment, contributing to increase the risk of environmental damage (Avancini et al., 2007; Aquino Azevedo, Ribeiro, Costa, & Amante, 2015).
Many functional properties, already evident in the fermentation of sour starch, may be related to organic acids present in the fermentation wastewater. Despite extensive studies of sour starch and its properties (Cárdenas & Buckle, 1980; Mestres & Rouau, 1997; Demiate, Barana, Cereda, & Wosiacki, 1999; Lacerda et al., 2005; Marcon et al., 2009; Aquino Azevedo, Ribeiro, Costa, & Amante, 2015) no studies were found considering the wastewater from the sour cassava starch fermentation process with antifungal potential.
Aiming to formulate a theoretical basis for the mechanisms of protection of post-harvest fruits, this work aims to verify how three species of fungi behave when subjected to different antimicrobial tests with the wastewater of the cassava starch fermentation (WCSF) of different origins and check if the antimicrobial action is related to the organic acids contained in this raw material. To the best our knowledge, there are no published reports of the use of WCSF as well as its probable antimicrobial functions that it may have.
The antimicrobial properties of WCSF were evaluated against B. cinerea, M. fructicola and C. gloeosporioides, which are among the most damaging fungi for a wide variety of fruits.
To study the antifungal capacity, in vitro, of the wastewaters of the cassava starch fermentation (WCSF), the mycelial growth rate index (MGRI) and the conidial germination of three important fungi species in the post-harvest of fruits and vegetables were measured. The experiments were conducted at the Plant-Pathogen Interaction Laboratory of the Crop Protection Department of the Faculty of Agronomy ‘Eliseu Maciel’ at the Federal University of Pelotas (Latitude: 31º 46' 19" S, Longitude: 52º 20' 33" W), Rio Grande do Sul, Brazil. WCSF chemical analysis were performed in Chemical and Food Science and Technology Departments of Federal University of Santa Catarina.
Material
WCSF from the main producing regions of the State of Santa Catarina, Brazil, were evaluated: WCSF1 - São João do Sul (Latitude: 29° 13' 32'' S, Longitude: 49° 48' 24'' W), WCSF2 – Ibirama (Latitude: 27° 3' 14'' S, Longitude: 49° 32' 36'' W) and WCSF3 – Agronômica (Latitude: 27° 15' 55'' S, Longitude: 49° 42' 40'' W). The collection followed a plan considering the geographical distribution of these companies in Santa Catarina State, which was assessed through visits and the collection of quantitative data on the production of this wastewater. The WCSFs were packed in hygienic packaging for transportation. Physicochemical analyzes (pH, titratable acidity and total soluble solids) were performed on arrival of the samples, which were stored in a freezer at -18 ± 1 °C. Before conducting antimicrobial tests, WCSFs were microfiltered on 0.22 µm polytetrafluoroethylene (PTFE) membranes (Allcrom, São Paulo, Brasil).
For the determination of the organic acids present in the samples and for the preparation of the acid solutions used to carry out the microbiological analyzes, the acids used were: lactic (purity 90%) and butyric (purity ≥ 99.5%) from Fluka (Buchs, Switzerland), acetic (purity ≥ 99.8%) and propionic (purity ≥ 99.8%) from Sigma-Aldrich (St. Louis, MO, USA). HPLC grade methanol was purchased from J. T. Barker (Mallinckrodt, Phillipsburg, NJ, USA). The WCSF used for all analyzes was distilled and purified on Simplicity equipment (resistivity 18.2 MΩ cm, Millipore, USA).
Methods
Identification of WCSFs' antimicrobial potential and organic acid patterns found in thismaterial, physicochemical characterization of WCSFs
The pH of the samples was determined by reading in a potentiometer (Quimis model Q 400 A, Diadema, São Paulo, Brazil), in triplicate. Total titratable acidity was determined using the official AOAC method 950.07 (AOAC, 2005).
Total solids (ST) were determined by gravimetry, in triplicate, with the weighing of 10 mL of the sample in Petri dishes, previously dried and tared, which were kept in an oven with forced air circulation (Fabbe, São Paulo) at 60 ° C, until constant weight.
The identification and quantification of organic acids present in WCSFs were performed according to the method described by Aquino, Azevedo, Ribeiro, Costa, and Amante (2015) using a high performance liquid chromatography system (HPLC) equipped with a quaternary pump and a degasser (LC-20AT), an automatic injector (SIL-10A), with a 20 µL loop, a diode arrays (SPD-M20A) operating at 220 nm wavelength, controlled by a workstation (CBM-20-A), with LC Solutions Software data acquisition system, all manufactured by Shimadzu Corporation (Kyoto, Japan).
As a stationary phase, a C18 column, Hichrom (150 x 4.6 mm - Berkshire, UK) was used. The mobile phases used, with an isocratic flow of 0.6 mL min-1, consisted of a solution of KH2PO4 0.02 mol L-1 with pH 2.88 ± 0.2 (adjusted with phosphoric acid) and methanol, 98:2 for lactic, acetic and propionic acids (Zotou et al., 2004), and 60:40 for butyric acid.
Preparation of solutions with organic acid standards for antifungal tests
To compare the effect of WCSFs with acid standards, solutions of lactic, acetic, propionic and butyric acids were prepared in the same concentrations found in WCSF, using the highest concentration of each acid among the studied WCSFs. To carry out the microbiological analyzes, stock solutions of all acids with a concentration of 10% were prepared. The mix solution was prepared from lactic, acetic, propionic and butyric acid standards in the proportion of 1: 1: 1: 1, also with a final concentration of 10%.
Obtaining and Isolating Pathogens
The antimicrobial activity of WSCFs was studied on strains of Botrytis cinerea, Monilinia fructicola and Colletotrichum gloeosporioides, isolated from symptomatic fruits of strawberry, peach and papaya, respectively.
2.2.5. Mycelial growth assay
The test was implanted in polyethylene Petri dishes, 60 mm in diameter, containing nutrient culture medium of PDA (Potato-dextrose-agar). 200 µL of the solutions under study were deposited on the culture medium and spread evenly with a Drigauski loop. To the center of each plate, a mycelial disc (8 mm) was transferred from a fungus colony with 10 days of growth in PDA medium. After inoculation, the plates, sealed with PVC film, were placed in a growth room and maintained at 24 °C, with a photoperiod of 12 hours of light, until the mycelium reached more than 2/3 of the plate.
The evaluation of mycelial growth was verified by daily measurements of the colony, obtained by the average of two diametrically opposed measurements, using a digital caliper, in millimeters (mm) and the mycelial growth rate index (MGRI) was calculated according to a formula adapted by (Araújo et al., 2008) (Equation 1).
MGRI = ∑ (D – Da) / N (1)
Where: MGRI = Mycelial growth rate index; D= actual mycelial median diameter; Da = anterior median diameter; N = number of days after peaking.
2.2.6. Lateral mycelial growth
The lateral mycelial growth test was performed using polyethylene Petri dishes, 60 mm in diameter, containing PDA, which were divided into two hemispheres. 100 µL of the solutions were spread evenly with a Drigauski loop on one side of the plate. A disc (8 mm) of mycelium, from fungus colonies with 10-days old, was transferred to the center of the plate hemisphere without solution. After inoculation, the plates were sealed with PVC film and kept in a growth room, at 24 ° C, with a 12-hour photoperiod. The lateral growth was evaluated visually every 24 hours, observing the area of the plate's hemisphere, containing the solutions under study, reached by the mycelia (0; 12.5; 37.5; 62.5 and 87.5%). The experiment was carried out twice for each fungus under study.
2.2.7. Conidial germination
For each treatment, a Petri dish with a fungal culture with 10 to 15 days old was used. In each plate, 5 mL of study sample and 10 µL of Tween 20 were deposited and, with a Drigauski loop, a suspension of conidia and mycelium was obtained. The suspensions were filtered through gauze and the amount of conidia was determined by counting in a Newbauer chamber, being adjusted to the concentration of 1 x 105 conidia mL-1.
90 mm polyethylene Petri dishes were divided into four quadrants and a 40 µL drop of the conidial suspension was deposited in triplicate in each quadrant. Then, each plate was placed open in a disinfected gerbox box, lined with paper moistened with sterile water to form a humid chamber. The suspensions were incubated at 27 °C, 100% RH and photoperiod of 12 hours. Fungal growth was halted 24 hours after incubation (time determined through preliminary tests) by the addition of 5 μL of lactophenol and 5 μL of Amann's Blue. Conidial germination was evaluated microscopically (200x) on 50 conidia per drop. Germinated conidia were considered as those with a sessile appressorium or a germ tube larger than the width of the conidia itself. The experiment was carried out twice for each fungus. Statistical Data were submitted to analysis of variance (ANOVA) and treatment means were compared based on Tukey's test (P <0.05). The data were analyzed using the Assistat 7.7 Beta (2015) software.
Characterization of the wastewater of the cassava starch fermentation process
The characteristics of the wastewater from the fermentation of cassava starch reflect the behavior of the starch granules subjected to the fermentation process. Although all starches are formed by amylose and amylopectin, the fermenting granules reflect the environmental conditions in which the cassava roots are grown, which can result in granules with large differences in size, compaction, enzymatic susceptibility, density, crystallinity pattern, and other properties inherent to the organization of this natural polymer (Aquino Azevedo, Ribeiro, Costa, & Amante, 2015). These factors can cause differences in the characteristics of soluble and suspended solids in the residual water from the fermentation of cassava starch, depending on the location where the roots are grown (Marcon et al., 2009). Wastewater of the three different origins studied differs mainly in terms of pH and soluble solids (Table 1).
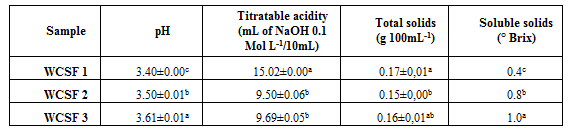
For each analysis performed, the values compare the averages of the different fermented cassava starch producers, averages followed by the same letter in the column do not differ significantly (p <0.05) by the Tukey Test.
Sources: WCSF 1 – São João do Sul; WCSF 2 – Ibirama; WCSF 3 – Agronômica.
Since the fermentation of cassava starch is a spontaneous fermentation, without the use of starter cultures, another important difference among the regions is the species of recurrent microorganisms, which can contribute to the variation of some parameters, especially the acidity, which corresponds to the specific pKa of the organic acids developed in the fermentation of cassava starch (Aquino Azevedo, Ribeiro, Costa, & Amante, 2015).
The total solids contained in the WCSF, probably consist of fragments of amylose and amylopectin molecules, in addition to organic acids and residual cells of microorganisms. Avancini et al., (2007) studied the WCSF obtained through a process carried out in the laboratory and in industry, regarding chemical, microbiological and toxicological characteristics. Authors identified that this wastewater (from industries or produced in laboratory) has a low amount of total solids, varying between 1,700 to 6,000 mg L-1 and, consequently, nutrients, presenting the following chemical characteristics: pH between 3.0 and 3.7; acid number from 21 to 68 mL of NaOH mol L-1 100 mL-1; 0.19 to 0.62 g of 100 mL-1 lactic acid.
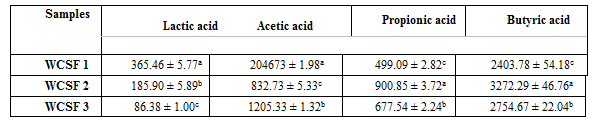
The concentrations of organic acids in the WCSFs analyzed were presented in the Table 2. In the WCSF 1 occurred the higher concentration of lactic and acetic acids while in the WCSF 2 occurred the higher concentration of propionic and butyric acids.
Identification of WCSF antimicrobial potential
Data of the conidia germination, mycelial growth and mycelial growth tests towards the antimicrobial compound (lateral growth) shown antimicrobial action of the WCSFs studied. Total inhibition of the B. cinerea and C. gloeosporioides conidia germination occurred to the three WCSFs (Table 3). On M. fructicola, WCSF 2 also inhibited the germination of 100% of the conidia, while in the treatments with WCSFs 1 and 3 occurred 5 and 2% of germination, respectively.
The effect of the standard acid on the three fungal (Table 3) showed that acetic, butyric acids and the mix were the most effective in inhibiting the germination of the conidia. In the WCSF 2 occurred the greater amount of butyric acid, when compared to other wastewaters, and it can be attributed to this acid the inhibitory action on the germination of three studied fungi by this WCSF.
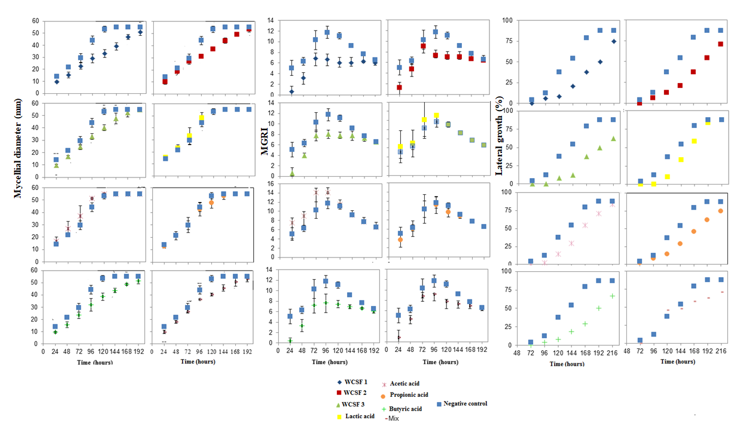
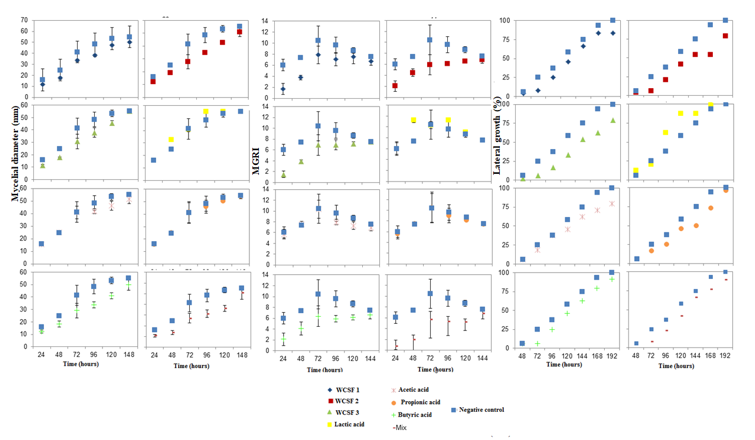
Lactic acid increased the conidia germination of B. cinerea, while on the other two fungi, both lactic and propionic acids significantly reduced conidia germination (Table 4). When the treatments with butyric acid and the mix were studied in isolation, similar responses to the treatments with wastewater for growth analysis and MGRI were verified. The butyric acid and mix solutions decreased the mycelium advance during the 192 h of incubation and the propionic acid, until the 144 h. Lactic and acetic acids did not show significant differences in relation to the negative control (Fig. 1).
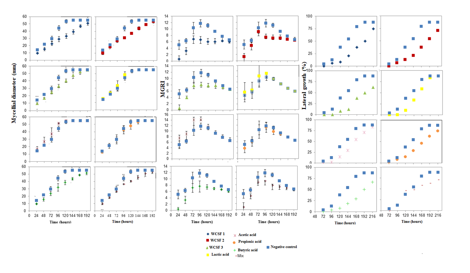
The mycelial growth of M. fructicola was more influenced by WCSF1 and butyric acid, and for the MGRI (Fig. 2), it was verified that, for the same treatments, there was a decrease in the growth speed, mainly in the first hours of incubation. For the lateral mycelial growth of M. fructicola towards treatments with WCSFs or butyric acid (Fig. 2) there was a significant decrease in relation to the negative control. Although less intense, there were also decreases in growth, in relation to the negative control, in treatments with acetic, lactic and propionic acids.
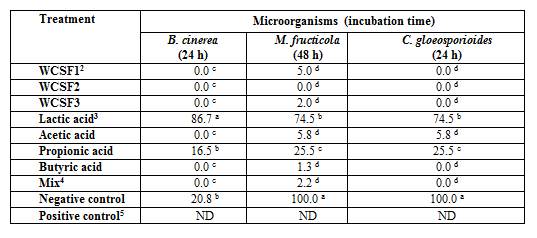
The values compare the averages of the different fermented cassava starch producers with the solutions made with the standards of organic acids in identical concentrations the highest among WCSFs; averages followed by the same letter in the column do not differ significantly (p <0.05) by Tukey test.
²WCSF1 - São João do Sul; WCSF2 - Ibirama; WCSF3 – Agronômica.
³ Standard solutions of the lactic, acetic, propionic and butyric acids on concentrations in mg L-1: 365.46; 2046.73; 900.85 e 3272.29, respectively.
4 Mix: solution of the mixture of lactic, acetic, propionic and butyric at the same concentrations.
5 Tebuconazol 100 ppm.
ND – non detected.
For the mycelial growth of C. gloeosporioides (Figurer. 3), the treatments had effects similar to those caused in B. cinerea. With the exception of butyric and mix acids, which showed less effectiveness in controlling the lateral growth of this fungus when compared to its action on B. cinerea (Figure 1).
Several approaches have been taken to study the effectiveness of the antimicrobial effects of acids on cells. These effects have been compared taking into account the pH, concentration, chain length, type and degree of branching to inhibit or kill a wide variety of microorganisms, nce these are the parameters that influence the growth or inhibition of many microorganisms (Hsiao & Siebert, 1999; Hassan, El-Kadi, Sand, & Hassan, 2015). With the addition of acids to foods, the pH decreases and, although fungi in general are the microorganisms most resistant to low pHs, the antimicrobial action depends on the acid species and characteristics such as concentration, type and buffer effect. So, it has been considered that the action of organic acids on fungi is more effective as a microbiostatic than as a microbiocide (Dolores, 2005).
The toxic effect of acetic acid on microorganisms is not only related to the decrease in pH, but also to the non-dissociated form of it that can cross the conidial membrane and penetrate the microbial cell and exert its toxic effect on cell protoplasm (Sholberg, Haag, Hocking, & Bedford, 2000). Several studies showed positive results in the use of acetic acid to control diseases in the post-harvest period. Acetic acid (2.5 µL mL-1) totally suppressed the germination of C. gloeosporioides in vitro (Cia, 2005). On apples, acetic acid was effective in in controlling B. cinerea and P. expansum without damage to the fruits (Sholberg & Gauce, 1995; Shouberg, Cliff, & Moyls, 2001). Acetic acid was also reported as an alternative to extend the shelf life of grapes by controlling B. cinerea Moyls, (Sholberg & Gaunce, 1996). Furthermore, use in the form of vinegar is also effective in reducing contamination by B. cinerea in cherries (Sholberg, 1998) and grapes (Sholberg, Reynolds, & Gaunce, 1996) and by M. fructicola in apricot and plums (Liu, Chu, & Zhou, 2002).
On the other hand, lactic acid in culture media increased the aflatoxin production by A. parasiticus, but the level of this mycotoxin was dependent on the pH of the medium (Luchese & Harrigan 1990; El-Gazzar, Rusul, & Marth, 1987). However, lactic acid is an excellent inhibitor of the development of bacterial cells at pH 5.0 (Woolford, 1975). However, it is not effective against yeasts and fungi.
The propionic and butyric acids reduced the growth of Aspergilus flavus and caused paralysis in the production of aflatoxin, being the propionic acid more efficient in controlling this fungus when compared to butyric acid (Ghosh & Häggblom 1985).
According to Adams (1999), organic acids have been considered responsible for the breakdown of amino acid metabolism, DNA synthesis and energy metabolism of microorganisms. Acids decrease intracellular pH and can cause changes in membrane permeability, blocking the substrate of the electron transport system. Weak lipophilic acids such as lactic, acetic or propionic are capable of permeating the cell membrane of microorganisms in their non-dissociated state and dissociate inside the cell, producing H + ions that lower the cell's pH. The cells react by eliminating the protons trying to keep the pH constant and this mechanism causes the energy expenditure to be higher, reducing the microbial cell growth. In turn, acid anions prevent DNA synthesis by preventing the protein from replicating (Choct, 2004).
With the results obtained, in this study, the antifungal activity of the wastewater of the fermentation of cassava starch was confirmed against the three fungi studied. This property can be attributed, in part, to some organic acids present in this material, taking into account that these had an evident antimicrobial action. In all analyzes performed with butyric acid, on the three fungi tested, there was a significant effect. The mix had an effect on B. cinerea and C. gloeosporioides. All treatments, with the exception of lactic acid, had an effect on lateral growth. Due to statistical differences, it was concluded that WCSFs have a broader and more effective inhibitory effect, when compared to the acids tested alone, and that this effect is possibly directly related to the concentration of the acids present, or to a synergistic effect between the acids, or even, to the existence of some other constituent in WCSF that potentiates this action, since such composition has not been fully characterized.
The results found in this study confirm the potential of this residue as a natural product with antimicrobial properties.
Authors declare any conflict of interest.
Declaration of conflict of interest
The authors declare no financial interests or personal relationships to influence the present study.
The authors thank CAPES (Coordination for the Improvement of Higher Education Personnel) – Juliane Mascarenhas Pereira Doctor fellowship. LJ Dallagnol is supported by fellowship CNPq (grant number 308149/2018-1). ER Amante was supported by fellowship CNPq (grant number 301688/2011-7 and 304741/2014-0).
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
