
Review Article | DOI: https://doi.org/10.31579/2642-9756/003
Clinical Department of Oncological Gynecology, Medical University of Lodz.
*Corresponding Author: Wójcik-Krowiranda Katarzyna, Clinical Department of Oncological Gynecology, Medical University of Lodz. E-mail: katarzyna.wojcik-krowiranda@umed.lodz.pl
Citation: Wójcik-Krowiranda Katarzyna, Szczepaniec Sylwia , Bieńkiewicz Andrzej, The Role of The βklotho Gene, Fgf21 and Fgfr1 in Cancerogenesis. J. Women Health Care and Issues, Doi:10.31579/2642-9756/003
Copyright: ©2018. Wójcik-Krowiranda Katarzyna. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 30 November 2018 | Accepted: 10 December 2018 | Published: 14 December 2018
Keywords: βklotho gene; fgf21; fgr1; endometrial cancer; cancerogenesis
Klotho was discovered in 1997 as an anti-aging gene that, when overexpressed, may extend the life span, but when it is disrupted, it may be a factor responsible for premature aging syndrome. The structure and the role of αKlotho and βKlotho genes from Klotho family in malignant tumors is described. The expression profile of the βKlotho gene is significantly different from the expression of the αKlotho gene. Analysis of Klotho expression in breast cancer, cervical cancer as well as endometrial cancer are discussed. The available data indicate the involvement of βKlotho in the neoplastic transformation of the endometrium. More advanced disease is related to negative expression of βKlotho gene. Fibroblast growth factors (FGFs) are a large family of proteins characterized by different functions in the cell development and metabolism. The FGF signaling is also associated with cancerogenesis. The relation between some FGF subfamilies and endometrial cancer clinical data is reported. The interaction between FGF subfamilies and the Klotho subfamily proteins acting as a co-receptor is stressed.
Disorders in signaling of the FGF / FGFR pathway have been confirmed in gynecology. It can be assumed that increased expression of FGF21 might be a suppressor factor in endometrial cancer. The FGF21 factor, like the βKlotho protein, achieves its biological effect via the FGFR1 receptor. High expression of the FGFR1 gene inhibits further tumor growth. FGFR1 has the potential to perform both a suppressor and promoter role in the oncogenesis process.
Significant progress in molecular biology caused a growing interest in searching for genetic background of malignant diseases. One of the most important features connected with neoplasmatic tumor growth is its immortality. The list of genes which play a role in the process of immortalisation is growing constantly. One of the most influencing genes responsible for the malignant cell life span is the Klotho family. The fibroblast growth factor (FGF) is another protein family involved in cell survival, proliferation and migration. The FGF family activation is moderated by the betaKlotho protein which plays a role of co-receptor necessary for FGF functioning in the cell. Thus, the relation between those two proteins in malignant tumors is still under investigation.
The structure of the gene and protein βKlotho
Klotho
Klotho was discovered in 1997 as an anti-aging gene that, when overexpressed, may extend the life span, but when it is disrupted, it may be a factor responsible for premature aging syndrome [1, 2]. The name of the gene and protein is derived from the Greek goddess Klotho, who along with the sisters Lachesis and Atropos spun the thread of human life and decided on its length [3, 4].
Klotho is composed of 1014 amino acids and has at its N-terminus signal sequences and the intra-membrane domain and short cytoplasmic domain at the C-terminus. The Klotho part of the trans membrane segment consists of two repeats (KL1 and KL2) that divides the homology sequence with β-glucosidase (Figure 1) [5].

Based on the similarity of the human DNA sequence to the sequence corresponding to αKlotho in mice, another 2 genes belonging to the Klotho family, βKlotho andIntrodusc γKlotho which is called KLPH (KL lactase phlorizin hydrolase) or LCTL (lactase-like protein) [6].
The βKlotho (KLB) gene encoding the monotopic transmembrane protein was discovered in 2000 as a homologue of the αKlotho gene (αKL) of mice (4). The human KLB gene, 44.68 kb in length, is located in the region 1 of the band 4 of the short chromosome 4 (4p14) and, similarly to the mouse, it consists of 5 exons and 4 introns. The mature transcript of this gene reaches a length of 6.08 kb [7-9].
The expression profile of the βKlotho gene is significantly different from the expression of the αKlotho gene. Studies in the mice KLB show that the expression of this gene is closely related to the metabolism of cholesterol and bile acids, and the presence of βKlotho protein in the pancreas, fat tissue and liver suggests that this protein may play a much wider role in metabolism (10), (11).In humans, KLB expression was detected mainly in the liver, fat tissue, glandular tissue of the breast and also in the spleen, testes, prostate, eyeball and brain [8] (Fig. 2).

βKlotho protein
The trans membrane protein βKlotho has a molecular mass of 119.8 kDa and is a homologue of the αKlotho protein. The level of amino acid compatibility between these proteins in humans is about 41.2% (86) As in the case of αKlotho protein, there are two repeated regions in the extracellular domain of βKlotho with structural similarity to one family of glycoside hydrolases (Figure 3) [4, 13, 14].

There are also important facts that distinguish βKlotho from its αKlotho homologue. Unlike αKlotho, previous studies have not shown that the βKlotho gene transcript undergoes an alternative assembly process that would result in the secretion of this protein. Also it does not seem that, as in the αKlotho protein, in the βKL2 region there was a short amino acid sequence KKRK, within which the protein would be proteolytically cleaved [13].
βKlotho
The Klotho gene is considered to be the youth gene (1) (2). The role of the Klotho gene transcript in the process of cancer is of great interest to scientists and has been studied so far in many malignant diseases. Immuno histochemical analysis of Klotho expression in breast tissue showed its high expression in healthy tissue and very low gene expression in breast cancer tissue. In tissue samples of breast cancer, high Klotho expression was correlated with a smaller tumor size and less mitotic activity. Detailed studies of breast cancer cells have shown that the Klotho gene as an inhibitor of the IGF-1 pathway (insulin-like growth factor-1) and also an activator of the FGF pathway (fibroblast growth factors) is a tumor suppressor [15].
Cervical cancer studies have shown that Klotho mRNA is absent in numerous samples of cancer tissue at advanced FIGO stages, but not in early pre-invasive (in situ) oncogenesis. After treatment, Klotho's expression returned to normal. Therefore, it was concluded that the reduction of Klotho expression may be highly advanced in cervical cancer.
The βKlotho gene from the Klotho family is also the object of interest of many scientists. The first data on this gene confirmed its role in the development of liver cancer. Then the study of the role of βKlotho in the progression of liver cancer gave divergent results [16, 17]. However, most studies suggest that βKlotho inhibits the proliferation of liver cancer cells. According to Poh et al. [16], increased expression of βKlotho and FGFR4 was detected in human liver cancer cells. A similar increase in expression was observed in HCC cell lines (hepatocellular carcinoma). This suggests that increased expression of KLB is associated with neoplastic transformation and progression of liver cancer. On the other hand, other data showed reduced KLB expression in HCC and HCC cell lines [17]. In addition, restoration of normal KLB expression in these cells inhibited their proliferation.
Until now, the level of KLB expression has been marked in various healthy and cancerous tissues (Figure 4).
The FGF21 gene is located in the region 5 of chromosome 19. Studies have shown that with obesity caused by both genetic factors and abnormal diet, the activity of the studied growth factor FGF21 induces a reduced level of glucose and insulin in the blood, a change in the lipid profile with a significant reduction in LDL and cholesterol levels.
the benefit of HDL and consequent weight loss through increased energy expenditure without the need to reduce meals [39]. In summary, the functions of FGF21 in many aspects is similar to the role of insulin (Table 2).

Receptors of FGFR fibroblast growth factors - structure and functions
Receptors for fibroblast growth factors (FGFRs) are monotopic transmembrane proteins with tyrosine kinase activity [41].\Data from the literature indicate that proteins belonging to the FGF15 / 19 subfamily (including FGF21) have an extremely low affinity for FGFR. To achieve the greatest ability to bind to and activate the receptor,the
FGF15 / 19 subfamily proteins must interact with the Klotho subfamily proteins acting as co-receptors [43-45].Co-operation of βKlotho with FGF21 and FGFR1Endocrine growth factors, among them FGF21, provide a biological cellular response depending on growth factor receptors (FGFR). The activity of FGF21 against FGFR is low even in the presence of heparin or heparan sulfate [46, 47]. To obtain a metabolic cellular effect, FGF21 requires the presence of βKlotho protein [48]. The impulse to transmit the FGF21 signal is a complicated process of creating a triple complex between FGFR1, βKlotho and FGF21.
FGFR1c may remain inactive despite the close presence of βKlotho in the absence of FGF21 [49]. Only the combination of FGF21 and βKlotho leads to the dimerization of FGFR1c in a manner allowing activation of the signal [50]. FGF21 and βKlotho act synergistically, increasing their chances of joining the receptor and thus obtaining a biological effect [50]. In addition, FGF21 itself contains amino acids at the N-end that determine its affinity for FGFR. The removal of 6 amino acid residues from the N-end of FGF21 significantly reduces its activity, whereas the removal of 8 amino acids abolishes it completely [51].Latest reports are increasingly related to genetic aspects in the epigenesis of malignant tumors. The life cycle of the cell is controlled at the genetic level. The general scheme of carcinogenesis is based on the initiation of a genetic error, its promotion and progression. The process of apoptosis prevents the fixation of genetic errors. All this determines the search for mechanisms responsible for cancer at the
genetic level of a single cell.The available data indicate the relationship between the expression of βKlotho genes, FGF21 and FGFR1, which at the cellular level, by regulating the life cycle of the cell and activation of metabolic pathways, may decide on the promotion of carcinogenesis or suppression of cancer. In the endometrial cancer, indirect indicators of malignancy seems to be: the clinical FIGO stage, the lymph node
metastases and the tumor differentiation (grading). The analysis of the relation between the expression value of the examined genes and the mentioned above factors in a representative cohorts, indicate the important role of the examined genes in the cancerogenesis of the endometrium.
FGF / FGFR pathway signaling disturbances
Various aberrations of FGFR signaling have been identified in tumors:
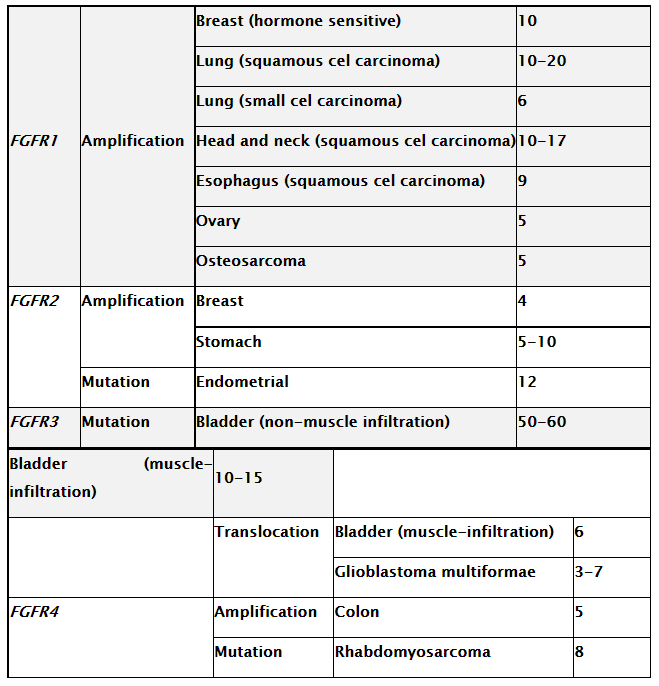
Disorders in signaling of the FGF / FGFR pathway have also been confirmed in gynecology. Fibroblast growth factors and their receptors play an important role in the proper development and functioning of the breast glands. Over recent years, it has been demonstrated that ectopic expression of FGF / FGFR family proteins may induce cancer transformation in breast cancer [53]. An important role in the progression of breast cancer by signaling initiated by fibroblast growth factors is attributed to FGFR1. Amplification of this gene was observed in 10% of patients with breast cancer. Of the 880 analyzed breast tumor specimens, the increased expression of FGFR1 mainly concerned HER2 negative (human epidermal growth factor receptor 2) patients over 50 years. It has also been shown that the amplification of the FGFR1 gene in women with estrogen receptor-ER breast cancer increases the risk of distant metastases and is associated with the progesterone receptor deficiency [54, 55].
In ovarian cancer, the gene encoding fibroblast growth factor 3 (FGF3) is amplified. Amplification of FGF3 was demonstrated in 20% of 136 examined cases. In addition, analysis of the results showed a significant correlation between the number of copies of the FGF3 gene and the FIGO stage classification [56]. Other reports show that FGF3 is expressed only in cancerous tumor tissue whereas it is not present in normal tissue [57]. A similar relationship was also demonstrated in FGF19. In ovarian cancer tissue,
amplification of the 5q31 region including the FGF1 gene has also been identified. Increased expression of FGF1 at the level of mRNA and protein was also correlated with a higher level of angiogenesis marker i.e. cluster31 differentiation antigen. These data may therefore suggest that FGF1 enhances the process of angiogenesis, which in turn may be the cause of worse prognosis of patients with FGF1 amplification [54, 58].
FGF21
Current reports are focused on deeper analyzes of FGF / FGFR pathway disorders. The growth factor of FGF21, which is the co-receptor of KLB [48], plays an important role in carbohydrate-lipid cell metabolism and has a positive effect on weight reduction [59, 60]. Both carbohydrate-lipid disorders and obesity are well known as a risk factors of the endometrial cancer. That is the reason why it is so important to elucidate the role of FGF21 in the endometrial cancer. The evaluation of FGF21 gene expression indicates that the high expression of the FGF21 gene (above 70.2) is accompanied by a strong dominance of the lowest clinical stage FIGO (stage 1). The lower values of gene expression are in turn accompanied by a significant increase in the share of higher (2 or 3) FIGO stages. It can therefore be assumed that increased expression of FGF21 might be a suppressor factor in endometrial cancer. Similar results have been obtained in the most recent studies of liver cancer [61], where it was observed that the initially increased expression of FGF21 is a defense and compensatory mechanism. However, in the late stage of changes taking place in the cell, the reduction of FGF21 expression may be associated with chronic liver disease, including neoplastic transformation. Considering the fact that obesity and diabetes associated with lipid disorders are common in patients with endometrial cancer, due to the associated reduction of FGF21 expression, these diseases are a direct risk factor for cancer development. Therefore, it can be assumed that patients with metabolic disorders are devoid of the protective effect of FGF21. As
is known, in most solid tumors, including endometrial cancer, there are numerous hypoxic outbreaks in which a low oxygen concentration may contribute to the reduction or even loss of FGF21 exposures leading to tumor development. Nevertheless, the mechanisms for reducing or losing FGF21 expression in endometrial cancer are not fully understood. In addition, FGF21 factor is the regulator of many metabolic processes in tissues with intensive expression of KLB [62]. Therefore, the further studies in this issue are needed.
In endometrial tumors, there was no statistically significant relationship between the presence of lymph node infiltration and FGF21 gene expression. In the majority of cases, the lymph nodes were not affected. The expression value of the FGF21 gene assumes a full range both in the group of patients with cancer and in the control group. On this basis, the local role of FGF21 can be attributed to the development of endometrial cancer without affecting lymph node metastases. This relation requires further analysis in a larger group, possibly also the evaluation of FGF21 expression in the lymph nodes themselves, in order to assess its local impact [63]. The histological differentiation of the tumor is of great prognostic importance. Studies on histological grading dependence on the FGF21 gene expression value did not show significant relationships. It is worth noting, however, that the expression of the FGF21 gene equal to 0 was not accompanied by a single case of endometrial cancer with a high tumor differentiation (G1) [63].
FGFR1
The FGF21 factor, like the βKlotho protein, achieves its biological effect via the FGFR1 receptor. Studies on the expression of FGFR1, depending on the clinical - histological factors of endometrial cancer, showed a strong relationship between the clinical and
pathological data of the tumor and the expression of FGFR1 [63].High values of FGFR1 gene expression occur only at the lowest FIGO stage. In the higher clinical stages (FIGO 2 and 3) the majority of cases with lower FGFR1 expression were observed. It can be assumed that high expression of the FGFR1 gene inhibits further tumor growth. This is probably due to the protective effects of βKlotho and FGF21 proteins, which reach their biological effect via the FGFR1 receptor. The influence of FGFR1 expression on the degree of histological differentiation (grading) is similar. Tumors of unfavorable maldifferentiation (G3) were usually accompanied by low expression of FGFR1. In the group with higher expression of the studied receptor, the incidence of G3 tumors decreased significantly. It also seems that the expression of FGFR1 is associated with the risk of metastasis of endometrial cancer in regional lymph nodes. Although the majority of data related to patients with lymph nodes were free of neoplastic infiltration, at low values of FGFR1 gene expression in 1/3 of the cases, the lymph nodes were occupied, whereas with higher gene expression, in almost 95% of cases the lymph nodes were not affected. The question, whether high expression of FGFR1 also has a protective effect on the formation of distant metastases requires further research. In the available literature, data evaluating the effect of FGFR1 expression on the development of endometrial cancer is limited [63]. FGFR1 expression was assessed in many tumors (Fig. 6),
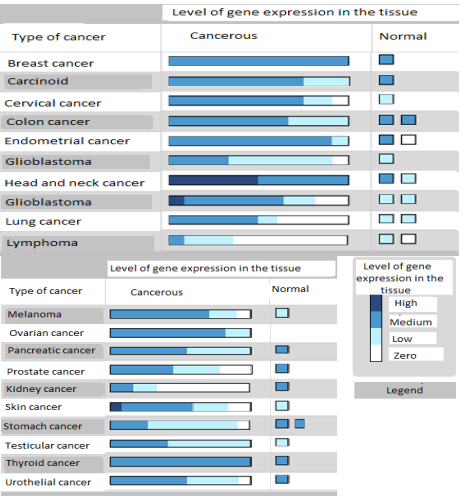
where significant differences in the expression of this gene in normal and tumor-altering tissue were demonstrated.
Recent studies have shown frequent focal amplification of FGFR1 in the non-small cell lung cancer cell line [65].It can be assumed that FGFR1 has the potential to perform both a suppressor and promoter role in the oncogenesis process depending on the cellular context that determines its ultimate function [66].Considering the histological structure of endometrial cancer, the above reports would confirm the results indicating the protective effect of FGFR1 on the development of glandular cancers, including endometrial cancer [63]. Confirmation of this dependence may be found in the other studies on prostate cancer where theFGFR1 is often overexpressed [65]. This mechanism is not fully understood as yet. Current reports indicate that over-expression of FGFR1 acts destructively on the subtle play of homeostasis between epithelial and mesenchymal cells [67]. In studies conducted on a prostate-specific mouse model, Acevedo and his colleagues directed the inducible FGFR1 gene to the glandular epithelium. After the activation of the FGFR1-specific epithelium, the epithelial to mesodermal transformation (EMT - epithelial to mesenchymal transition) and increased adenocarcinoma formation were observed. Deactivation of FGFR1 in the early stage of tumor development leads to complete tumor regression, suggesting the role of this gene in both initiation and progression of prostate cancer. When the inactivation of FGFR1 occurred in the late stage of neoplastic transformation, proliferation and progression of the tumor were limited, but no complete regression of the tumor was observed. In this way, differences in susceptibility to FGFR1 inhibition at different stages of the disease were demonstrated, indicating "susceptibility windows" [67]. This window can be regulated by other factors which verification could reveal new therapeutic options.The EMT process itself is significant in the development of cancer, especially due to the emergence of distant recurrences and metastases. Cells created in the EMT process have enormous regenerative capacities (compared to the regenerative capacity of stem cells) and migrate to distant parts of the body, where they revert to epithelial cells, form a cluster and metastatic tumor. Thanks to this mechanism, the number of cells necessary to produce a tumor from a million is reduced to 10,000 [68].
Studies on the role of FGFR1 in the formation of distant metastases were conducted in prostate cancer mice model. FGFR1 inactivation in prostate cells, resulted in smaller tumors developed that contained discrete foci of cells still expressing FGFR1, indicating a focal isolation of tumor foci to protect against gene excision. In addition, tumors with a confirmed deletion of FGFR1 showed an increase in outbreaks of well-differentiated cancer. On the other hand, tumors that avoided the removal of FGFR1 mainly showed a poorly differentiated phenotype. According to these phenotypes, mice carrying the zero-expression allele of FGFR1 survived much longer than those without the FGFR1 cut. Interestingly, all metastases, mainly absent at the zero FGFR1 allele, showed high expression of FGFR1 and the neuroendocrine phenotype, regardless of the expression status of FGFR1 in primary tumors. Together, these results suggest the critical role of allowing ectopic FGFR1 signaling in the formation of prostate tumors and, in particular, metastasis [69]. In conclusion, it should be noted that, βklotho, FGF21, FGFR1 genes and the proteins encoded by them play an important role in the endometrial tumors formation. Further research on the participation of these factors in carcinogenesis processes in the endometrial mucosa will be a starting point for the development of effective methods for modifying the expression of these genes and for influencing the synthesis of their encoded proteins. This will allow the implementation of molecularly
dedicated therapeutic processes in accordance to currently developing research trends in oncology.
List of Abbreviations
3T3-L1 – line of mouse fat cells
αKL – gene α Klotho
cDNA -complementary DNA
CYP1A1 - cytochrome P- 450 1A1
DEPC -Diethylpyrocarbonate
ER – α Estrogen receptor alpha
ER- β Estrogen receptor beta
FGF -fibroblast growth factor
FGFR -fibroblast growth factor receptor
FIGO -fr. Fédération Internationale de Gynécologie et d'Obstétrique
G1, G2, G3 - grading
HCC - hepatocellular carcinoma
HBS -heparan sulfate binding site
HPRT1 -hypoxanthine-guanine phosphoribosyltransferase
HS - heparan sulfate
IGF – 1 -insulin-like growth factor 1
KRAS -Kirsten rat sarcoma viral oncogene
KLB – βKlotho gene
KLPH - KL lactase phlorizin hydrolase
LCTL -lactase-like protein
MTHFR- methylenetetrahydrofolate reductase
PR A -progesterone receptor
PR B -progesterone receptor
PTEN -phosphatase and tensin homolog
SHGB -sex hormone binding globulin
SERM -selective estrogen receptor modulator
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
