
Review Article | DOI: https://doi.org/10.31579/2692-9406/105
International Higher School of Medicine, Intergelpo str, 1F, Bishkek, 720 040, Kyrgyzstan.
*Corresponding Author: Abyt Ibraimov, International Higher School of Medicine, Intergelpo str, 1F, Bishkek, 720 040, Kyrgyzstan.
Citation: A Ibraimov, S Akhunbaev, O Uzakov. (2022). The Missing Link in the Human Thermoregulation Systems. Biomedical Research and Clinical Reviews. 6(4); DOI: 10.31579/2692-9406/105
Copyright: © 2022 Abyt Ibraimov, This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 07 February 2022 | Accepted: 22 February 2022 | Published: 28 February 2022
Keywords: cell thermoregulation; human thermoregulation; condensed chromatin; chromosomal heterochromatin; human body heat conductivity
The study of human thermoregulation is one of the most developed areas of modern physiology and medicine. This is due to the important role of temperature in normal and pathological conditions. Directly or indirectly almost all systems and organs are involved in maintaining of relatively constant temperature in the body, since their normal functioning is closely related to the temperature of the internal environment (temperature homeostasis).
Currently, the existence of two systems of thermoregulation is recognized: a physiological organ-based system with a center in the hypothalamus and a molecular system, which manifests itself in the form of the production of proteins (heat shock- and cold shock proteins) and by activation of RNA thermometers in response to heat or cold shock.
Based on studies of the variability of chromosomal heterochromatin regions in the genome of human populations permanently living in various climatic and geographical conditions of Eurasia and Africa, as well as individuals well adapted to the extreme conditions of the high altitudes of the Pamirs and Tien-Shan (mountaineers) and the Far North (oil drillers of the Yamal Peninsula, Eastern Siberia), we came to the conclusion that, apparently, there is a third system of thermoregulation at the cellular level (cell thermoregulation). By cell thermoregulation, we mean the removal of excess heat from the nucleus of an interphase cell in order to avoid the harmful effects of high temperature on the complex molecular processes occurring in the nucleoplasm.
We believe that some intracellular formations (a dense layer of condensed chromatin around the nucleus, nucleoli, chromocenters and other membraneless bodies) serve as a structural basis for the removal of heat from the cell nucleus, because the material basis of all these temporary structures in the interphase nucleus is the chromosomal heterochromatin regions.
In this review, we discuss a set of data, which indicate that cell thermoregulation, may be a missing link between organism and molecular level in maintaining temperature homeostasis in the human body.
The study of human thermoregulation is one of the most developed areas of modern physiology and medicine. This is due to the important role of temperature in normal and pathological conditions. Directly or indirectly almost all systems and organs are involved in maintaining of relatively constant temperature in the body, since their normal functioning is closely related to the temperature of the internal environment (temperature homeostasis).
Normally, human life as a homoeothermic organism can take place in a narrow temperature range, and this condition is ensured by complex thermoregulation systems. As is known, thermoregulation is a set of physiological processes aimed at maintaining the relative constancy of temperature in the body under conditions of changing ambient temperature by regulating heat production and heat transfer. Thermoregulation is aimed at preventing violations of the body's thermal balance or at restoring it, if such violations have already occurred. Thermoregulation is carried out by the neuro-humoral way and changes in the level of metabolism, which leads to a change in the formation of heat in the body.
As is known, the main sources of heat (heat production) in the body are exothermic reactions of oxidation of proteins, fats, carbohydrates, ATP hydrolysis, as well as muscle contraction. However, in order to maintain a relatively constant temperature in the body, it is necessary to remove excess heat in a timely and efficient manner using heat transfer mechanisms. In thermo physiology, four main mechanisms of heat transfer are recognized: evaporation, heat conduction, heat radiation and convection.
The efficiency of heat transfer of the human body is exceptionally high. Biophysical calculations show that a violation of these mechanisms, even in a human at rest, would lead to an increase in his body temperature within an hour to 37.5 °C, and after 6 hours to 46-48 °C, when irreversible destruction of protein structures begins. Violation of the ratio of these processes leads to a change in body temperature [1]. At the same time, the question remains unclear what is the material basis such a high heat-removing abilities of the human body, apart from its ability to excessive sweating and hairless skin?
Currently, the existence of two systems of thermoregulation is recognized: a physiological organ-based system with a center in the hypothalamus and a molecular system, which manifests itself in the form of the production of proteins (heat shock- and cold shock proteins) and by activation of RNA thermometers in response to heat or cold shock [2].
However, in our opinion, another important question remains open; namely, how does heat transfer occur at the level of individual cells? It is considered that four recognized mechanisms of heat transfer (evaporation, heat conduction, heat radiation and convection) are necessary and sufficient conditions so that dangerous temperature increases do not occur in the human body. Since, ultimately, the main source of heat is the processes occurring in cells, and then there must be some mechanisms and means capable of removing excess thermal energy beyond its limits. To be more precise, the question can be posed as follows: are there mechanisms and corresponding structures that ensure maintaining temperature homeostasis at the cell level. If so, what are the structural components of cells involved in the removal of excess heat beyond its limits? In other words, is there thermoregulation at the cell level, and what are the roles of evaporation, heat conduction, heat radiation and convection in removing excess heat from the cell? Obviously, the roles of evaporation, heat radiation and convection in cell thermoregulation should be very limited. Therefore, the cells could solve the problem of heat dissipation mainly through changes in the thermal conductivity of some of its structural elements [3].
Undoubtedly, the main biochemical processes responsible for heat production in the human body are carried out in the cytoplasm and they are well studied. It is taken for granted that the excess heat produced in the cytoplasm is removed without problems into the intercellular space and from there, with the help of a circulation system, is removed outside the human body.
However, the question arises, what is the role of the nucleus in thermoregulation? After all, no less complex biochemical and molecular biological processes occur in the nucleoplasm than in the cytoplasm, which are also accompanied by the release of heat. In particular, internal (repair, recombination, rearrangement, modification, restriction) and external (replication, transcription, packaging, organized movement) molecular activity of chromosomes is accompanied, among other things, by the release of heat. If the nucleus temperature begins to exceed the temperature of the cytoplasm, there is a need for the removal of excess heat outside the nucleoplasm. From the point of view of physics, the nucleus has two possibilities: to increase its volume and/or to increase the heat conductivity of the nuclear envelope. Although the first possibility is limited for known biological reasons, the second possibility seems more promising, in the sense that there is no physical or physiological restriction for this. Apparently, Nature has long solved this problem, otherwise how could, for example, homoeothermic organisms arise and safely exist. Therefore, our task is to find these heat-dissipating intracellular structures, to study their nature and mechanisms of work [4].
Based on studies of the variability of chromosomal heterochromatin regions in the genome of human populations permanently living in various climatic and geographical conditions of Eurasia and Africa, as well as individuals well adapted to the extreme conditions of the high altitudes of the Pamirs and Tien-Shan (mountaineers) and the Far North (oil drillers of the Yamal Peninsula, Eastern Siberia), we came to the conclusion that, apparently, there is a third system of thermoregulation at the cellular level [5].
We believe that in the study of human thermoregulation system, two links are missing: thermoregulation at the level of individual cells and the heat conductivity factor of the human body. By cell thermoregulation, we mean the removal of excess heat from the nucleus of an interphase cell into the intercellular space in order to avoid the harmful effects of high temperature on the complex molecular processes occurring in the nucleoplasm [3,5-7].
As is known, heat conductivity is the process of transferring internal energy from more heated parts of the body to less heated bodies, carried out by chaotically moving micro particles (atoms, molecules, electrons, etc.), which leads to equalization of body temperature. Heat conductivity is not accompanied by the transfer of matter. This type of internal energy transfer is typical for both solids and liquids and gases. Therefore, there is nothing unexpected in the idea that the human body should have some heat conductivity.
However, from the literature we could not find information about the results of special studies aimed at estimates of the magnitude of the heat conductivity of the human body in vivo. Apparently, this is due to the well-known fact that the human body by its nature, including density, is deeply heterogeneous. Nevertheless, in recent years we have managed to collect the first experimental data on the existence of variability in the level of human body heat conductivity (BHC) in the population. Moreover, it turned out that individuals differ from each other in the BHC depending on their gender, age and climatic and geographical features of their places of permanent residence [8].
Many years ago we have postulated that the condensed chromatin (CC) in interphase nucleus may be concerned with intracellular thermoregulation. CC, being the most densely packed material, appears to have the highest heat conductivity in the interphase cell [3]. By CC we mean: (1) tissue-specific condensed euchromatin (particular portions of euchromatin condense and differentiate as part of a tissue-specific transcriptional control system); (2) the facultative heterochromatin and (3) constitutive heterochromatin [10].
In our opinion, all that is known about chromosomal heterochromatin regions (HRs), interphase nuclei, and redundant DNA of higher eukaryotes does not contradict the idea of a possible heat conduction role of CC in the cell nucleus. We include the following points:
(1). The periphery of cell nucleus is occupied by heterochromatin, which is closely associated with the lamina and the inner nuclear membrane; and the nucleoli are surrounded by dense CC [11-19] (Fig.1).
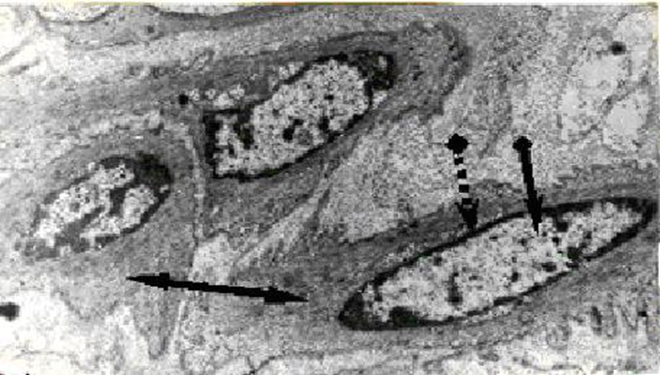
(2) Redundant DNA in most eukaryotic organisms is complexed with proteins in highly compact structures designated as CC. Chromosomal HRs are a particular case of differential packaging of the redundant repetitive DNAs [20].
(3) The centromeres of chromosomes with nucleolus organizing regions (NORs) are consistently associated with nucleoli and in the human genome all rDNA loci are embedded in constitutive heterochromatin. Genes of 18S, 28S, and 5S RNA, tRNA, and mRNA of histones are located on these sites.
(4) The chromosomal G, Q, and R-bands are absent in plants and are always present in the chromosomes of higher vertebrates (reptiles, birds, and mammals). In the case of invertebrates, fish, and amphibians, it is difficult to reveal these bands [13,21]. The difficulties of revealing G, Q and R bands in plants, insects, other invertebrates, fish, and amphibians are frequently explained as methodological difficulties. But we believe that they are not connected to the reproducibility of techniques of differential staining and reflect the true state of affairs [3,5-7].
(5) Chromosomal HRs of autosomes 1, 9 and 16, which do not contain ribosomal cistrons always contacts with the nucleolus [22].
(6) Variability of the amount of chromosomal HRs, not only within but also between species occurs with no visible phenotypic effect [23].
(7) At the later stage of evolution of the ancestors of the three higher primates (Homo sapiens, Pan troglodytes, and Gorilla gorilla) a new type of constitutive heterochromatin—Q-heterochromatin— appeared [24-26].
(8) Chromosomal Q-heterochromatin regions (Q-HRs) exist in the genome of the three higher primates, but broad quantitative variability exists only in human populations [27-42].
(9) Changes in the amount of Q-HRs in the human populations have a tendency to decrease from low to high geographical latitudes and from low to high altitudes [33-41].
(10) Since wide quantitative variability of chromosomal Q-HRs was detected only in human population but alimentary obesity, alcoholism and drug addiction relate to the group of "purely human being pathology" we thought that it would be quite logically to study peculiarities of BHC of such diseased individuals. It was established that among patients, drug addicts had the highest BHC, then alcoholics and individuals suffer from obesity [42-45].
As known any serious scientific hypothesis has to be verifiable. But what experiment or natural system can be offered to verify that hypotheses? It might be reassuring, for example, if someone managed to show that when change the temperature in the human cell culture above or below 37 °C, the speed and amount of heat transfer from the nucleus to the cytoplasm depends on the amount of chromosomal HRs.
In some sense, Nature seems to have done us a favor and conducted part of work for us. For example in the process of evolution, redundant repeated DNAs in the genome of higher eukaryotes appeared as CC at the periphery of the interphase nucleus: in plants as C-heterochromatin, in the chromosomes of higher vertebrates, in addition to C-heterochromatin, G+ and Q+ bands appeared. Internal temperature environments of animals are more stable than in plants. Amphibians and reptiles have a body temperature partly below the ambient one, but birds and mammals control temperature homeostasis within very narrow limits. The appearance of the stable internal environment allowed these animals to counteract variations in temperature. Birds and mammals developed organ-based control of thermoregulation and have a neural ‘thermostat’ situated in the hypothalamic region of the brain. The order Carnivora is quite a unique group among homoeothermic animals as regards dense l ayer CC around the nucleus. Identification of heterochromatin regions by means of the existing technique is extremely difficult [46,47]. It is possible that their known mode of life requires lower heat conductivity inside cells, and eventually the whole body.
Since the proposed idea is based on cell phenomena, from our point of view, nature “found” a very simple and effective solution: it increased the level of human BHC through compression the density of CC around layer of the nucleus with chromosomal HRs. Below are the results of some observations that indirectly indicate the reality of cell thermoregulation in humans.
Of course, the cell thermoregulation (CT) hypothesis needs to be checked in vivo at the cellular level, but so far such experiments are still impossible. It turned out that it is even more difficult to directly assess the level of human body heat conductivity (BHC) due to the complete lack of any experience in this regard. In particular, it is still not possible to develop a method to accurately measure the human BHC in the way that it is done for homogeneous nonliving objects by physicists. Nevertheless, we have checked this hypothesis at the level of the human organism assuming that CT is the material basis for heat conductivity of the cellular part of the body (for details see [8,47].
The aim of these studies were to look for a possible link between the amount of Q-HRs in the genome and human BHC, that is, whether a random set of chromosomal Q-HRs in the human genome can determine the level of thermal conductivity of his body? In other words, whether there is a connection between the number of chromosomal Q-HRs and its level of BHC?
The amount of chromosomal Q-HRs in the genome of human populations is usually estimated by two quantitative characteristics: the distribution of individuals according to the number of Q-HRs in their karyotype in population and the derivative of this distribution, an important population characteristic, is the mean number of Q-HRs per individual.
In thermo physics, it is customary to measure the heat conductivity of solids (for example, metal) by the calorimetric method. Heat energy is transferred through a metal rod, the ends of which are placed in calorimeters with water, and temperatures are measured at T1 and T2 (T1 > T2). To assess the heat conductivity, the amount of heat transferred and the time required to measure the thermal conductivity coefficient of a given metal rod are determined. Obviously, this method of measuring heat conductivity is unacceptable for studying the human body for both technical and ethical reasons.
By trial and error, we determined the areas of the body and the mode of thermal load (the creation of a temperature gradient between the body and the water bath with a thermostatic device), which would allow us to assess the level of human BHC (high, medium and low). Our experience has shown that the most informative data are: (a) the time of the onset of the temperature peak on the surface of the right palm during a controlled heat load; (b) the temperature difference (T) between the surface of the right palm and the oral cavity, and (c) the T of the right palm at the time of maximum temperature increase (for more details, see [7,8]).
Table 1 shows the relationship between the number of chromosomal Q-HRs and the rate of reaction of the body to the controlled thermal load, which was determined by the time (in minutes) to the occurrence of peak temperature on the surface of the right palm.
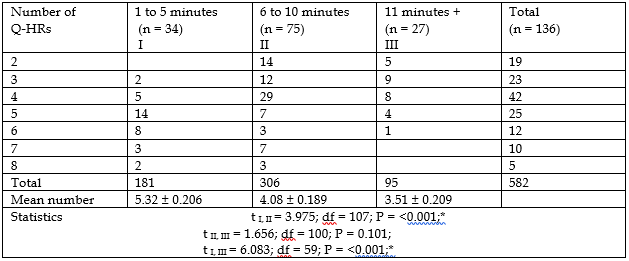
Table 1 show that there is a statistically significant relationship between the number of chromosomal Q-HRs in the human genome and the reaction of the body to the controlled thermal load. In individuals whose genome contains more amount of chromosomal Q-HRs, the peak temperature on the surface of the right palm occurs in the first five minutes of the thermal load, and vice versa.
The relationship between the amount Q-HRs and the temperature difference between the right palm and the oral cavity at rest is shown in Table 2.
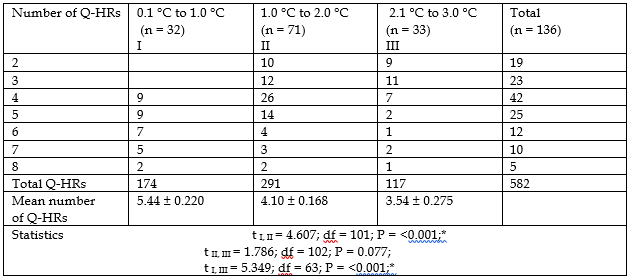
As can be seen in Table 2, the more chromosomal Q-HRs, the smaller the T difference between the oral cavity and the right palm, and vice versa.
Table 3 shows the relationship between the number of chromosomal Q-HRs and the value of T of the right palm at the moment peak temperature occurs during the controlled thermal load.
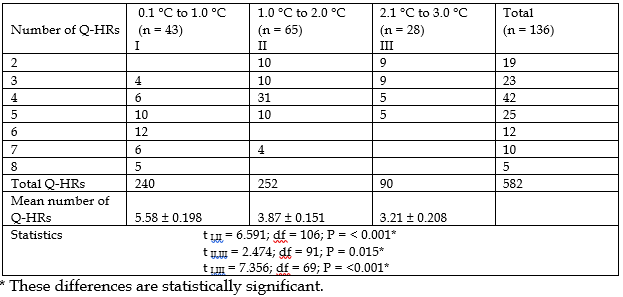
As shown in the table, there is a statistically significant relationship between the number of Q-HRs and the value of T of the right palm at the moment of peak temperature occurrence – in individuals with a large amount of Q-heterochromatin, the T of the right palm rises less, and vice versa.
How do we interpret these data? We believe that the time to occurrence of peak temperature on the right palm reflects the rate of heat conductivity. If the peak temperature on the surface of the palm occurs in the first five minutes after the thermal load, then such an individual is considered to have a high BHC, and vice versa. In other words, a person with a high BHC conducts heat through the body quicker and dissipates any excessive through the body shell quicker.
A statistically significant relationship between the amount of Q-heterochromatin and the T difference between the oral and hand at rest may also characterize the heat conducting ability of the body – the smaller the T difference, the higher the BHC, and vice versa. We believe that a smaller T difference between the oral cavity and the palm reflects the high heat conductivity of the body, so that such an organism can successfully avoid overheating.
These data show that there are differences in the BHC between individuals in a population. In particular, we were able to show that individuals in a population differ significantly from each other in terms of BHC level. We also found that the level of BHC is affected by sex, age, and the climatic and geographical features of the individual’s place of origin. However, the BHC level is not affected by weight, height, arterial pressure, pulse rate, and respiration [47]. In other words, there are some parallels beween the distribution of chromosomal Q-HRs (see above) and the variability of BHC at the level of human populations.
Which of the existing biological phenomena underpin this wide human BHC variability in human populations? The first thing that comes to our mind is, of course, the basal metabolic rate, which is a well-known factor in physiology. However, it is also known that the core temperature of those living in the tropics lies within a similar range to those dwelling in the Arctic regions. Apart from that, the basal metabolic rate is influenced by such factors as height, weight, body constitution, pulse rate, and environmental temperature, which contradicts our data [47,48].
In terms of possible genetic (materials) factors, the most appropriate is the amount of chromosomal heterochromatin (C- and Q-heterochromatin) in human genome. But we suppose that the packing density (compactization) of CC is basically connected with the amount of chromosomal Q-heterochromatin in the nucleus [5]. The point is that human populations do not differ significantly in the quantity of C-heterochromatin in their genome [49,50]. A wide quantitative variability at the level of human populations is found only in the amount of chromosomal Q-heterochromatin. Thus, it is apparent that human BHC variability depends primarily on the amount of chromosomal Q-heterochromatin in the genome. As the amount of chromosomal heterochromatin does not change in ontogenesis, it is possible that the level of BHC may have a constitutional character, like the color of skin, eye shape, bodily constitution, height, and other innate physical human peculiarities.
The essence of the issue under discussion can be reduced to the following, why is it impossible to completely reject the possibility of the existence of a third link in the thermoregulation system in humans and animals? There are questions that have not yet received a rational scientific explanation. Below are some of them and as we believe, are related to heat conductivity.
As is known, homoeothermy is a later acquisition of the evolution of the animal world. By the way, there is no scientific explanation for the existence of warm-bloodedness, except for the fact that homoeothermic animals are able to maintain a constant within 2 ° C with relatively wide fluctuations in ambient temperature. Science does not yet know the origin of homoeothermic animals and the material substrate on the basis of which homoeothermic organisms arose in the process of evolution [4]. For example, penguins, partridges, ducks and other birds do not freeze in winter because the temperature of their paws may differ from the body temperature by more than 30 degrees. It is possible that the heat conductivity is essential here (for example, the low heat conductivity of the paws could greatly reduce heat transfer).
The unique ability of man, the only species that has managed to inhabit all the land of the Earth, including such extreme areas as Far North and high altitudes, remains not fully elucidated. It is not only the scale of the territory that is striking here, but the time (around 30,000 - 50,000 years) that it took for a human to adapt to a variety of climatogeographic conditions. At the same time, human was and remains a single tropical biological species. In addition, the genetic mechanisms that formed the basis of human adaptation and origin are not known [7,51].
Since wide quantitative variability of chromosomal Q-HRs was detected only in human population but alimentary obesity, alcoholism and drug addiction relate to the group of "purely human being pathology" we thought that it would be quite logically to study peculiarities of BHC of such diseased individuals. It was established that among patients, drug addicts had the highest BHC, then alcoholics and individuals suffer from obesity [43,44].
The example from the modern sport life can better illustrate our understanding of BHC role. More and more countries situated at low latitudes have started taking part in the world sport movement. The most notable in this process is that, natives of this region achieve great success in sports, requiring (in addition to other factors) effective heat-loss (football, professional boxing and marathon race). While sportsmen from high latitudes prevail in water and winter sports and also in mountaineering. Since southerners’ bodies, as we think, have relatively high heat conductivity it is not surprising, that they are successful in sports, which require effective heat-loss. Indeed, a sportsman with high heat conductivity cannot make much progress in water sports due to the fact that their body cools rapidly. However, this sportsman can be more successful in sports which require effective heat-loss [52].
It is known that with age the number of women in a population begins to prevail over men. It is possible that a certain advantage of women is explained by their relative resistance, as compared with men, to cold and stress, hunger and even loss of blood because they have less heat conductivity body. In order to be convinced of the relative resistance of women to cold and stress, we shall give several known examples: a) pearl divers in Korea are exclusively women - “ama” [53]; b) women succeed best in swimming across the cold water of La Manche [53]; c) during the period of the Leningrad blockade during the World War II about 80% of the women survived despite the fact that being in the rear they had a lesser access to food. Мы неоднократно убедились, что уровень BHC у женщин достоверно ниже, чем у мужчин [7].
The essence of the hypothesis of cell thermoregulation is to reduce the temperature difference between the nucleus and the cytoplasm, when the nucleus temperature becomes higher than the temperature of the cytoplasm. The nucleus, in contrast to the cytoplasm, cannot conduct heat directly in the extracellular space, from where the heat is taken by the circulating flow of sap, lymph and blood. Thus, the nucleus can conduct heat only in the cytoplasm. The role of the circulatory systems (CS) has not been discussed here in maintaining temperature homeostasis. The thing is the CS cannot influence directly the temperature inside the cells, as they are linked with the CS indirectly through the intercellular space. The only exception may be endothelial cells lining the inner surface of blood vessels. Thus, the CS influence on inner cellular temperature homeostasis is limited. That is why it seems that the problem of maintaining the inner cellular temperature homeostasis is solved by cells themselves, and we call it the cell thermoregulation [7,54].
As we suppose, during his evolution man, possibly owing to wide (broad) quantitative variability of chromosomal Q-HRs, had an additional and very flexible tool to ensure more effective thermoregulation, allowing him to master almost all the oykumene [55-58]. H. sapiens is not only devoid of a more or less large anatomic structure, but also has no protein or enzyme that has no analogue in the animal world. The fundamental structural characteristic of man is the presence of chromosomal Q-HRs in its genome which he has inherited together with the chimpanzee and the gorilla – from one common ancestor. Apparently, the physiological thermoregulation functions relatively independently from cell thermoregulation as evolutionally new adaptive system. From our point of view, cell thermoregulation can be the missing link, which should fill the "gap" between the thermoregulation systems, functioning at the molecular and the physiological levels. It is likely that we faced with physiological problem which is a new and alien for classical courses of physiology (for more details see [7]).
In essence the idea proposed here is reduced to the evolution of the genome structure and the physiology of the whole organism in higher eukaryotes going in parallel to counteract changes of temperature in the ambient environment for more effective preservation of constancy of temperature of the internal cell environment. The outcomes of such a parallel evolution were: (1) the appearance of different kinds of condensed chromatin (C- and Q-heterochromatin, G+ and Q+ bands, sex chromatin body; B-chromosomes, inactivation one of the X chromosome in mammals) at a cell level and (2) formation at an organism level of a complex organ-based physiological system of thermoregulation.
And finally, here we discussed the problem of thermoregulation only in relation to humans. In fact, they concern all homoeothermic organisms. Due to the fact that the possibility of thermoregulation at the cell level was indicated by research data on the wide variability of chromosomal Q-HRs, inherent only to the H. sapiens, we deliberately limited ourselves to discussing the problems of thermoregulation on the example of the human body.
I apologize to that author whose work is not cited or is cited only through reviews. The reason for this is only the space limitations of the publication.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
