
Review Article | DOI: https://doi.org/10.31579/2766-2314/008
AGH - Univ. of Science and Technology, Cracow, Poland.
*Corresponding Author: Maciej Pawlikowski. AGH - Univ. of Science and Technology, Cracow, Poland.
Citation: Pawlikowski M. (2020) The CO2 level as a factor stimulating angiogenesis Phenomena of tissue regeneration and destruction J, Biotechnology and Bioprocessing 1(2); DOI:10.31579/2766-2314/008
Copyright: © 2020, Maciej Pawlikowski, This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 30 October 2020 | Accepted: 02 November 2020 | Published: 17 November 2020
Keywords: CO2 level; angiogenesis; tissue regeneration
The regeneration of vessels and their development occur in places of their destruction. This phenomenon suggests that the destruction process itself stimulates vascular development and entails tissue healing and self-repair. Understanding the phenomena of self-repair is extremely important in the process of restoring tissues and organs to the proper functioning of individual organs and the entire body. Therefore, the recognition of angiogenesis processes seems to be the key in the treatment of many diseases.
Many years of research on blood vessels, their development and mineralization [15, 17, 18, 19, 24, 25, 26, 27,28, 29, 30, 31, 33], bone mineralization, demineralization and the process of bone healing [12, 13, 32], as well as the phenomena of tumor mineralization [32, 16, 20, 21, 25, 27], have all provided a lot of observations on angiogenesis. The presented publication is a summary of many years of early research.
Views on the role of endothelium in angiogenesis are widely published [1, 2, 7, and 10]. Various substances have been proposed as a factor in the development of arteries, including vitamin D3. An important role in the development of angiogenesis is also attributed to perisytes, i.e. unorganized stem cells present in the vicinity of blood vessels [4]. The literature in many publications also reports on the formation of new vessels in neoplastic tissues [14. 32, 34, 36, 39]. The literature also provides examples of phenomena that inhibit the development of angiogenesis [3], often based on their computer modeling [5, 11].
Literature information and the results of own research suggest a similar cause of angiogenesis in various cases of vascular formation. This view will be presented below.
Collateral circulation
In vessels blocked by substances crystallizing in them [18, 26], e.g. coronary vessels, the level of CO2 increases. This is the result of hypoxia, i.e. a change in the ratio between the oxygen level and the CO2 level at the point of blockage. Therefore, in the area of the artery blockage the oxygen level is low compared to the CO2 level, which increases as a result of the death of myocardial cells. Consequently, this phenomenon leads to the formation of easily dissociating carbonic acid (H2CO3), formed from the local CO2 and water present in the tissues. As a result of this, the pH of the environment drops below 7.0. The angiogenesis itself, developing at the site of arterial blockage, can lead to the formation of collateral circulation (Photo 1).

In the case of neoplastic tumors, the newly formed concentrations of cells form numerous blood vessels that develop from both primary and secondary vessels (Photo 2). The rate at which these blood vessels form is often slower than the rate at which new cells are formed. That results in an increase in non-discharged CO2 causing local acidification of the environment.

The phenomena described in this publication and their interpretation are based on studies of damaged arterial walls. It was noted that a damaged (for various reasons) artery wall may heal itself. The endothelial surface, which proliferates as a result of angiogenesis, forms particular structures that in further evolution lead to the formation of a new artery (Photo 3).

Endothelium is the site of many biochemical phenomena. One of the most important is the synthesis of prostacyclins, prostaglandins and thromboxanes, which are secreted into the blood and regulate its properties, including coagulability and the ability to aggregate. Research shows that individual elements of the cell membrane of intima have different functions (Figure 1).
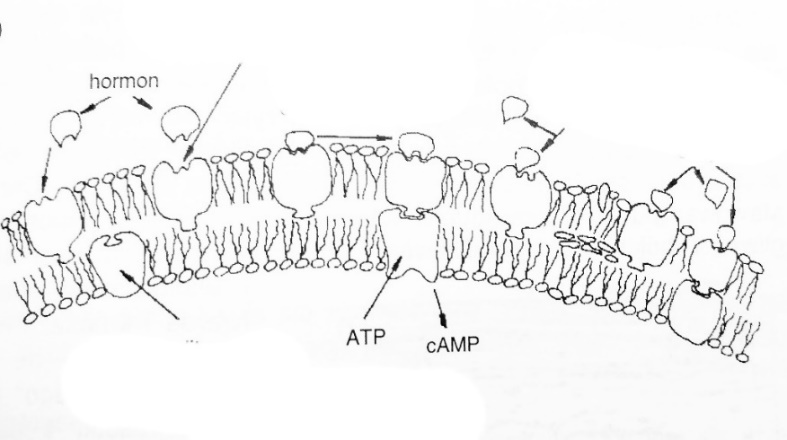
Concentrations of cholesterol crystallizing in and on the artery wall can inhibit blood flow. This causes local increase in CO2 content in relation to the content of oxygen, which is not supplied by the blocked artery. The phenomenon causes local CO2 to react with water, leading to the formation of carbonic acid (H2CO3). It dissociates easily and breaks down into free protons (H+) and CO32- carbonate groups. The above phenomena lead to a decrease in the local pH and development of acidic
environment that is aggressive towards the artery, where the flowing blood has a pH of 7.2-7.3. The result is that the acidic environment damages the endothelium of the artery.
The place of damage (destruction) of the artery endothelium is the site where interatomic bonds of its biological structures are broken. Due to broken bonds in the area of destruction an electric field appears, i.e. a place where free protons, carbonate groups etc. can be attached. Thus, the site of endothelial damage is the center in which the construction of new endothelium and consequently new vessel takes place (Figure 2).
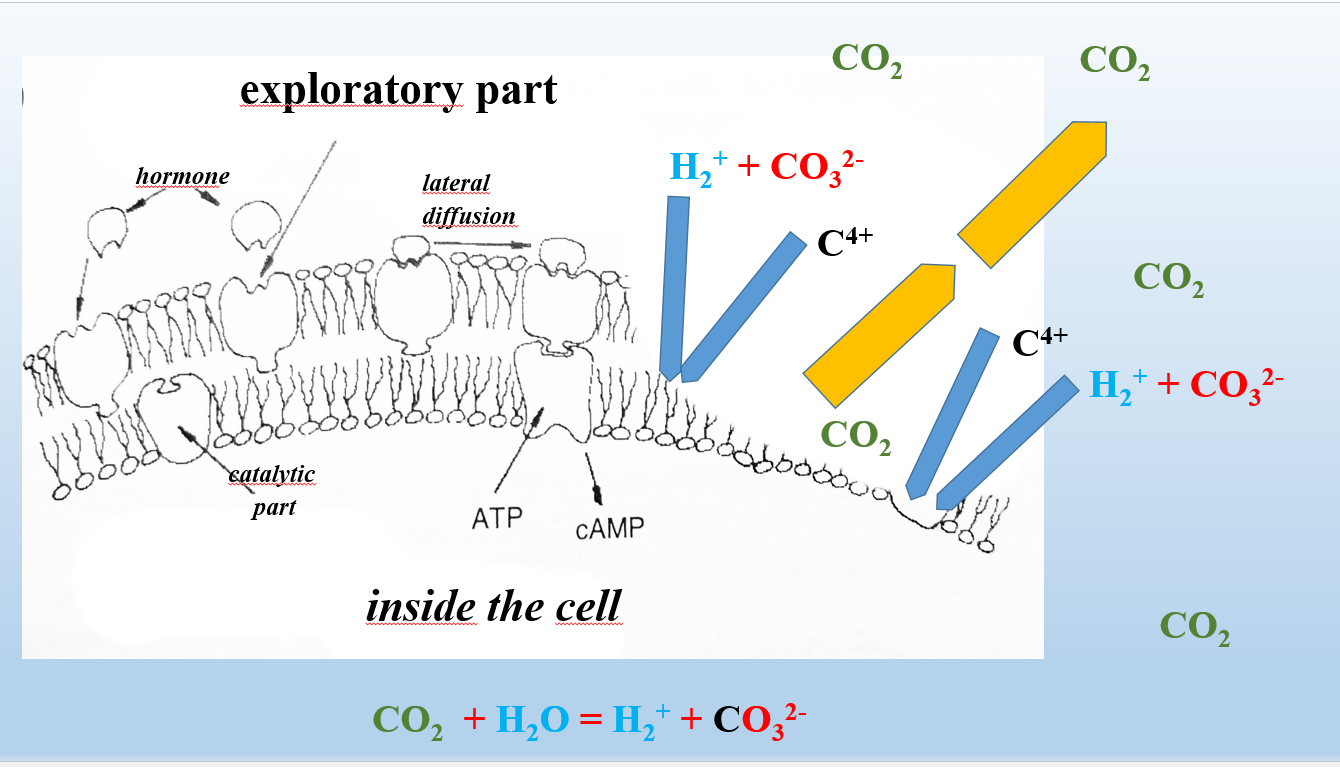
Protons and carbon from the CO32- groups build new biological structures at the site of damage to the endothelial cell membrane (blue arrows). The newly formed vessel (artery) proliferates in the direction shown by the yellow arrows.
In the case of angiogenesis in cancerous tumors, the phenomenon is similar. However, the cause of the local increase in the
level of CO2 is different than in the case of artery blockage. Here, the reason is the huge number of cells that multiply, each of them producing carbon dioxide in its life processes. The locally existing blood vessels are not able to evacuate the excess, which results in an increase in its quantity. The next stages of the phenomenon of angiogenesis generation and development proceed as described above, in the case of arterial blockage (Figure 3).

Damaged endothelium proliferates using “local material” in the form of protons and carbon to rebuild and initiate a new vessel.
While the cause of angiogenesis in bone fracture healing is different, it actually comes down to the phenomena described above for blocked arteries and tumors. In this case, there is also a local increase in the CO2 level. It is caused by the rupture of blood vessels during the fracture. The environment at the fracture site (especially in the fracture gap) contains an increased amount of CO2. This is caused by the inability
to drain it through damaged vessels. Increased amount of carbon dioxide leads to the formation of dissociating carbonic acid and the phenomenon continues as described for blocked arteries and tumors.
Due to the fracture, the arteries are damaged mainly in the form of ruptures. Thus, the damage occurs at the tips of the broken arteries, and that is where angiogenesis begins. As a result, the proliferation of newly formed vessels in the fracture zone takes place from the bone fracture to the bone fracture (Figure 4). The process of angiogenesis itself is similar to that in tumors and blocked vessels.

The phenomenon of new artery generation in the artery endothelium at the site of vessel rupture. Material for the newly developing artery is collected from the local environment (blue arrows). The direction of growth of the newly formed vessel runs roughly from one bone fragment to the other (yellow arrows).
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
