
Review | DOI: https://doi.org/10.31579/2690-1919/224
* Retired from Georg-August University, Institute of Microbiology and Genetics, Goettingen/Germany.
*Corresponding Author: Frank Mayer, Am Hohen Tore 4a, 38118 Braunschweig/Germany.
Citation: Frank Mayer. (2022). The Bacterial Cytoskeleton based on Bacterial Translation Elongation Factor EF-Tu: Novel Insights. J Clinical Research and Reports, 10(2); DOI:10.31579/2690-1919/224
Copyright: © 2022, Frank Mayer. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 19 December 2021 | Accepted: 30 December 2021 | Published: 10 January 2022
Keywords: bacterial cytoskeleton; elongation factor EF-Tu; occurrence; origin; formation; multi-resistance; pathogenic bacteria; target for antibiotics
Bacteria possess an EF-Tu-based cytoskeleton.This article presents a short review. A number of questions which are not discussed in the former publications can be asked, such as: all bacteria possess a ribosomal protein synthesis system and, hence, also EF-Tu. EF-Tu is produced in an amount that is higher than the need for a function as translation elogation factor in ribsomal protein synthesis. This article tries to answer the question regarding the surplus of EF-Tu: formation of a "cell-wide web" by self-assembly as a feafure that stabilizes cell integrity.
An additional question can be asked: what is the origin of this bacterial cytoskeleton? This article contains a speculation on this topic. A third question regards the'ntteructjon of ribosomes in the process of protemsynthesis: does the EF-Tu protein move to the ribosome, or does the ribosome move to the EF-Tu intergated in a fibril of the bacterial cytoskeleton? The former publication depicts electron micrographs which show colocalizatton of botth entities. EF-Tu is an example for aprotein with two independent functions: participation in the ribosomal protein synthesis as a kanslation elongation factor, and component of a bacterial cytoskeleton. This situation can open up a discussion ofthe sequence of events and states of early cells during evolution.
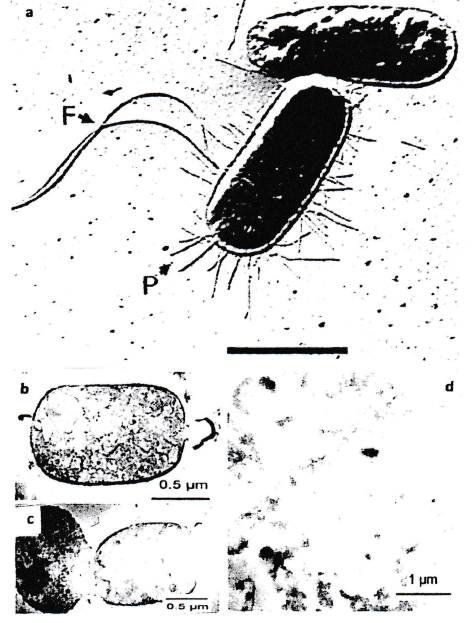
In former publications [1-6] and (Figure 1), the structural organization of the EF-Tu-based bacterial cytoskeleton is described. It is based on the surprising discovery of its existence [7,9].
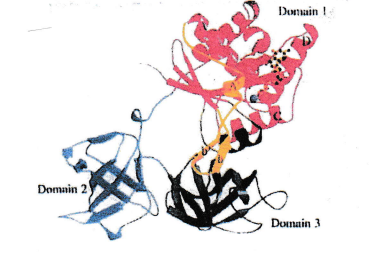
The protein exhibits three domains [5].
A bulge at domain 3 fits into a cleft in domain 2. This situation allows chain formation (fibrils, s. below) [6].
After all, the elongation factor EF-Tu is known for its important role in the bacterial ribosomal protein synthesis. In a former publication [10] it was stated that Ef-Tu in bacterial cells is produced in much higher numbers as compared to the need of its role in protein synthesis. No explanations were offered for the reason of this fact. In addition, in this former publication it was reported that EF-Tu can form fibrils consisting of many EF-Tu molecules bound to each other by non-covalent binding. Further implications of this observation were not discussed.
The idea that bacteria contain a cytoskeleton did come up by a surprising phenomenon: bacteria did maintain their typical shape and size even after removal of all layers of the cell wall and of its cytoplasmic membrane [11]. At this time publications such as [8] did report that bacteria possess a cytoskeleton consisting of proteins related to actin. Experiments with various kinds of antibodies, using immuno electron microscopic investigations, did not show binding of this labelling system to a layer close to the inner face of the cytoplasmic membrane when anti-actin antibodies were used. This layer close to the inner face of the cytoplasmic membrane mentioned above had to consist of other proteins. The properties of EF-Tu described above did lead to the conclusion that it may be EF-Tu that constitutes this layer. Immuno electron microscopy using anti-EF-Tu antibodies did give a convincing labelling result. In addition, positive labelling was also observed at other sites of the bacterial cells when thin-sectioning was applied for the labelling experiments.
Inspection of the results of an x-rax investigation of the EF-Tu protein [5] did give an explanation: EF-Tu is a 3-domain protein. The formation of fibrils by EF-Tu mentioned above could easily be explained by an interaction of an exposed distinct site (a bulge) on domain 3 with a distinct site (a cleft) exposed on domain 2 of the protein. In the meantime, the exact amino acid sequences of these sites are known [12].
This interpretation was supported by electron microscopic images obtained during investigations of fibrils isolated from artificially lysed bacteria, and of fibrils created, by polymerization, using isolated EF-Tu proteins (Figure 2).
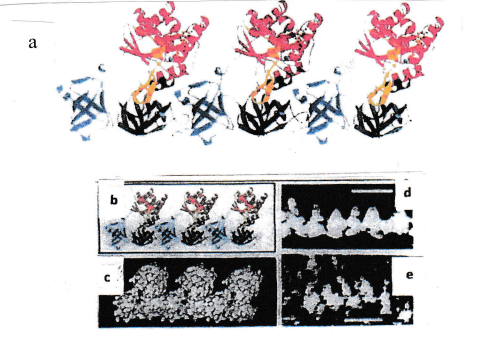
A further set of experiments was performed: isolated domain 3 protein was mixed with intact full-size EF-Tu. As soon as the domain 3 protein was located at the end of a fibril, the domain 3 had no binding partner (because domain 2 was missing) and, hence, fibril formation was hindered (Figures 3 and 4).
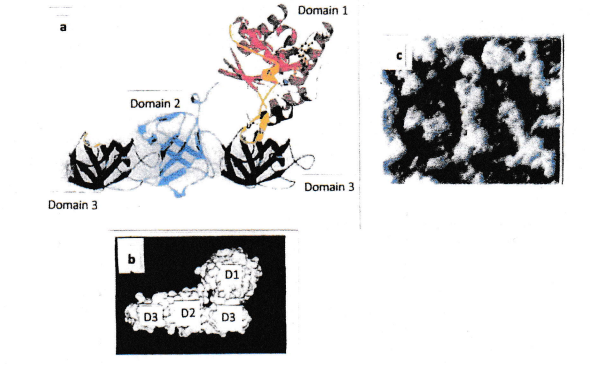
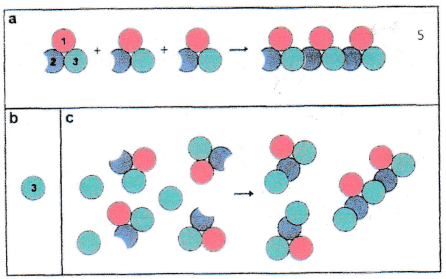
This observation was the starting point for investigations related to the use of cytoskeleton damage by a new class of antibiotics [13]. (Figure 5) depicts the result of the application of domain 3 for destruction of the bacterial cytoskeleton.
Note the increasing time-dependent damage to cell integrity .
Original electron micrographs by Frank Mayer
Novel insights
Existence of EF-Tu and EF-Tu-based cytoskeletons in all bacteria?
EF-Tu is a very old and conservative protein. Obviously, nature had no reason for selection of sequence variations in this very important protein.
As bacterial ribosomal protein synthesis is not missing in any bacterium, it is concluded that all bacteria possess EF-Tu protein molecules, probably synthesized in heigh numbers. This allows the conclusion that all bacteria possess an EF-Tu-related cytoskeleton. The surplus of EF-Tu in the bacterial cell (s.above) can allow that the cytoskeleton is formed by self-assembly: the individual EF-Tu proteins find their partners for the formation of fibrils by this process, with the energy needed for movement of the protein molcules and the binding process originating from Brownian movement. As EF-Tu protein is present in high numbers, a self-assembly would continuously take place during the entire growth phase of the bacteria. The consequence is an increase of fibrils in the bacterial cell (a “cell-wide web“, CWW). One could observe that, very often, a colocalization of ribosomes with cytoskeletal fibrils was evident. Hence, the question is: do the ribosomes their job (participation in protein synthesis) in close contact with EF-Tu proteins organized in fibrils? As the domain 1 of EF-Tu (Figure 1) extends towards the cytosol, and as the active center for elongation is located in domain 1, ribosomes close to an EF-Tu fibril might have easy access to domain 1. This would mean that EF-Tu protein is not moving to the ribosome to perform elongation, but that the ribosomes move to the EF-Tu proteins in the fibril. The observed colocalizaion of ribosomes and EF-Tu fibrils might support this view.
These described facts and conclusions indicate that bacterial cells, including multi-resistant pathogens, are targets for application of EF-Tu-based antibiotics [13].
By the way: development of resistance against EF-Tu or “truncated” EF-Tu may not be expected. After all, EF-Tu is an important component of ribosomal protein synthesis [9]. Resistance against EF-Tu would most probably damage the protein synthesis system, followed by death of the bacterial cell.
One more important aspect should be considered. Possession of a bacterial cytoskeleon is the common property of all bacteria. This is a feature that allows successful treatment of multi-resistant bacteria with the EF-Tu-based antibiotic even when all other antibiotics fail.
Speculations on the origin of EF-Tu and on its role in bacteria for cell stability
Protein synthesis is the key for living systems. In this context the overall stabilization of cell-like spheres by a cytoskeletal system may be of secondary importance. Hence, it might be reasonable to assume that precursors of modern EF-Tu systems active in ribosomal protein synthesis did not contain binding sites for fibril formation for cytoskeletons. Such a property might have evolved later. It should be kept in mind that the active center for the process of elongation of the growing protein is located in domain 1. The roles of domains 2 and 3 in this process (if present at all in early states of evolution) remain unknown (s.above, today’s functional role of these two domains). It can be speculated that two domains, now with binding sites, were added later in evolutioa. The EF-Tu-based cytoskeleton might be the primordial cytoskeleton in bacteria.
This view is supported by the fact that a different kind of bacterial cytoskeleton (MreB protein, related to actin of higher organisms) was found, by labelling experiments, to colocalize with the EF-Tu-based cytoskeleton [14,15]. It was even shown that the EF-Tu-based cytoskeleton plays a steering role regarding the placement of MerB elements in an EF-Tu – MreB complex, and in the placing of these complexes in the bacterial cell. It can be speculated - regarding evolution - that the EF-Tu-based cytoskeleton was first. The MreB cytoskeleton evolved later, and it then found its place in a complex of these two cytoskeletons by the assistance and steering role of the EF-Tu-based cytoskeleton components. As mentioned above, EF-Tu is a very old and conservative kind of protein. This fact supports the view that EF-Tu was developed very early in evolution, prior to many other proteins in the evolving bacterial cells. The cells could now use the potential for protein synthesis and optimization of protein function.
This line of interpretation would imply that the EF-Tu-based bacterial cytoskeleton is, as mentioned above, the primordial cytoskeleon. This view would add a new aspect to the discussion on the origin of bacterial cytoskeletons and, in general, of cytoskeletons.
Former publications (references) describing the structural and functional features of a bacterial cytoskeleton based on the translation elongation factor EF-Tu did not consider some aspects that might need further explanation and discussion. The present article contains a short review of the earlier basic findings, but novel insights should be added. An example is the formation of the cytoskeleton. It appears reasonable to assume that it is the process of self-assembly that is the mode how structural elements (full-size EF-Tu protein molecules) are forming chains (fibrils) that are building up networks stabilizing the bacterial cell. Formation of a “cell-wide web” is caused by the fact that synthesis of EF-Tu produces more EF-Tu that is needed as an elongation factor. The surplus assembles to fibrils which contribute to cell stability. This stability can be reduced by insertion of “truncated” EF-Tu (only domain 3 of the EF-Tu protein) into the growing cytoskeleton.
This situation was used for the design of a novel kind of antibacterial agent. This agent is synthesized by the ribosomal protein synthesis system inside the pathogenic bacterial cell as soon as the genetic information coding for domain 3 of EF-Tu has been transferred into the cell, and translation is started. This approach can be described as a kind of induced suicide": the antibacterial agent that kills the cell is produced inside the cell that had produced it.
A further example of a novel insight may be the questions around the origin of the first bacterial cytoskeleton. It was shown that MreB, an action-related bacterial protein, also forms cytoskeletal elements. It was also described that it is the Ef-Tu-based cytoskeleton that organizes the sterical organization of complexes that contain MreB and EF-Tu.
EF-Tu is a very old and conservative protein that did not have substantial sequence alterations during evolution. Together with the colocalization of EF-Tu and MreB, this fact supports the view that an EF-Tu-based cytoskeleton might be the first cytoskeleton in the first cells formed during the early stages of evolution. This would mean that the EF-Tu-based bacterial cytoskeleton is the primordial cytoskeleton. The active center in EF-Tu for the function of elongation is located in domain I of the protein, supported by few features located in domain 2 of the protein. Are domain 2 and domain 3 younger than domain 1?
Discussion is open
An additional question could be the interaction of ribosomes in the process of protein synthesis with fibrils of the EF-Tu protein. In one of the earlier publications [11] a colocalization of both entities was observed. Is soluble EF-Tu in the cell moving to the ribosome, or fist the ribosome moving to the EF-Tu frbnl? Are there any steric hindrances?
To sum up: The existence of the Ef-Tu-based antibacterial agent is an example for new insights and ideas in a field of research that is important the fight against multi-resistant pathogenic bacteria.
This work was carried at the Institute of Microbiology and Genetics, Georg-August-University Goettingn/Germany. Supported by Volkswagenstiftung and Deutsche Forschungsgemeinschaft.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
