
Review Article | DOI: https://doi.org/10.31579/2690-4861/746
1 Department of Neurology, Hospital “Sant’Antonio Abate” Trapani, Italy.
2 Retired Professor of Human Anatomy. University of Palermo. Palermo, Italy.
3 Department of Medicine and Surgery of Acceptance and Emergency. Emergency Room (MCAU) of the Villa Sofia-Cervello Hospital in Palermo, Italy.
*Corresponding Author: Farina Elvira, Retired Professor of Human Anatomy. University of Palermo. Palermo, Italy.
Citation: Lipari Alessio, Farina Elvira, Lipari Luana, (2025), Somatotopic Organization of Cranial Nerves of the Medulla Oblongata: IX, X, XI, XII, International Journal of Clinical Case Reports and Reviews, 25(1); DOI:10.31579/2690-4861/746
Copyright: © 2025, Farina Elvira. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 07 March 2025 | Accepted: 24 March 2025 | Published: 04 April 2025
Keywords: nuclei of medulla oblongata; cranial nerves of medulla oblongata; somatotopy
Stemming from our previous studies regarding somatotopy of the spinal cord, oculomotor, trigeminal complexes and facial nuclei we examine the nuclei of origin of cranial nerve of the medulla oblongata to understand some neurological clinical and surgical aspects.
The cranial nerves, twelve pairs, emerge from inferior surface of the brain and are designed by a name and by Roman ordinal numerals (I-XII) from rostral position to caudal position. The cranial nerves differ from the spinal nerves; in fact, all spinal nerves are mixed, originating from the union of a ventral motor root and a dorsal sensory root, while the cranial nerves differ from each other in structure and function. The cranial nerves some are only motor, some only sensory and some are both sensory and motor. In relation to the parts of the brain the cranial nerves are divided into: telencephalic nerve, I or olfactory nerve, which originates from the olfactory neurons in the olfactory epithelium and extends to olfactory bulb; diencephalic nerve, II or optic nerve, which originates from ganglion neurons of the retina of the eye and extends to the lateral geniculate body; mesencephalic nerves III, IV, V; pontine nerves VI, V, VII, VIII; medullary nerves V, IX, X, XI, XII. An Overview of the cranial nerves was reported by Porras-Gallo and coll [1].
Galen (129—216 circa) was the first to distinguish nerves, tendons and ligaments and, also, to distinguish the “spinal nerves" originating from the spinal cord and "encephalic o cranial nerves" originating from the brain and indicates numerating seven pairs of brain nerves: 1, Optic; 2; Oculomotor; 3-4, Trigeminal; 5, Facial and Auditory; 6, Glossopharyngeal-Vagus-Accessory; 7, Hypoglossus.
Acuña and coll [2] report that Mondino de' Liuzzi (1265–1326) [3], Italian anatomist, wrote the first modern anatomical text Anothomia (1316) based on the dissection of human cadavers maintaining the classification of 7 pairs of cranial nerves in: I, optic; II, oculomotor; III, abducens; IV, trigeminal; V, facial; VI, vagus, VII, glossopharyngeal. Eustachio (1552) [5], Colombo (1559) [5] and Falloppio [6] indicated I nerve the optic nerve and II nerve the oculomotor nerve. that was differentiated in three nerves: oculomotor, abducens, trochlear, but Falloppio [6] indicated: the II pair the oculomotor nerve; the IV pair the abducens nerve, added a new VIII nerve the trochlear and, also, indicated the III cranial nerve (trigeminal) subdivided in three branches ophthalmic, superior maxillary and inferior maxillary or mandibular.
However, except for Thomas Willis’s [7] version, these proposals were unsuccessful. Bartholin Gaspar father (1611), as Porzionato [8] reports, introduced the nervus olfactorius as the I cranial nerve and Willis [7] in his Cerebri anatome (1664) was the first to list the olfactory tracts as the I pair cranial nerve and proposed a new classification composed of nine pairs cranial nerves; the I, II, III, IV, V, and VI cranial nerves were those we recognize today; Willis included VII pair the facial and vestibulocochlear nerves, VIII nerve encompassed the glossopharyngeal and vagus nerves, and the cranial root of the accessory nerve; the IX nerve consisted of the hypoglossal nerve and the spinal root of the accessory nerve [9]. The Willis’s formula by its popularity was consolidated and remained in place for more than 100 years until 1778, when Sömmerring [10] in his Doctoral Dissertation formulated the current classification composed of 12 cranial pairs although it during the nineteenth century undergoes many disputes.
The above-mentioned classifications of cranial nerves are represented in the following table.
| Nerves | Galen | Vesalius 1543 | Colombo 1559 | Falloppio 1561 | Willis 1664 | Bidloo 1685 | Sommering 1778 |
0/N(Nulla)/ I Terminal (near lamina terminalis) | Ayers H (1919) numbered it “cranial nerve I”, considering a total of 14 cranial nerves including also the septal—today vomeronasal—nerve. (Ayers: Vertebrate cephalogenesiss. IV. Transformation of the anterior end of the head, resulting in the formation of the nose. J Comp Neurol 30:322–342). Demsky and Schwanzel 1987 numbered it “cranial nerve 0” because it was rostral to the other twelve nerves. (Demsky LS and Schwanzel-Fukuda M: The terminal Nerve: Nervus Terminalis, Structure, Function and Evolution. Annals of the New York Academy of Sciences. Vol. 519). | ||||||
| I | X | X | X | X | Tract | Tract | Olfactory |
| II | 1° p. | 1° p. | 1° p. | 1° p. | 2° p. | 2° p. | Optic |
| III | 2° p. | 2° p. | 2° p. | 2° p. | 3° p. | 3° p. | Oculomotor |
| IV | X | X | 9° p. | 8° p. | 4° p. | 4°p. nerve pathetici | Trochlear |
| V | 3° p. 4° p. | 3° p. 4° p. | 3° p. 4° p. | 3° p. | 5° p. Trifacial | 5° p. nerve gustatorii | Trigeminus |
| VI | X | X | 8° p. | 4° p. | 6° p. | 6° p. Nervi timidi | Abducens |
| VII | 5° p. | 5° p. | 5° p. | 5° p. | 7° p. Facial (portio dura) acoustic (portio mollis). | 7° p. | Facial
|
| VIII | Vestibulo-cochlear | ||||||
| IX | 6° p. | 6° p. | 6° p. | 6° p. | 8° p. | 8° p. | Glosso-pharyngeal |
| X | 9° p. | Vagus | |||||
| XI | 10° p. | Spinal Accessory | |||||
| XII | 7° p. | 7° p. | 7° p. | 7° p. | 9° p. | 11° p. | Hypoglossal |
| I Cervical | 10° p. | ||||||
Table 1: Classifications of cranial nerve from Galen to Sömmerring
Recent literature data report some errors in the cranial nerves concerning their emergence and classification to be corrected. Corrales [11,12] evidenced that all illustrations of contemporary atlases analyzed characteristically show CNs VI, VII, and VIII all emerging from the pontomedullary groove as originally depicted by Sömmerring [12]. Differently the MRI findings show that the exit location of CN VI was caudal to the CN VII/VIII complex in 93% of cases and, also, multiple MR images analyzed coupled with microsurgical experience of this anatomical region indicate that CN VII enters the pons caudal to CN VIII. Revising a universally recognized numbering scheme would certainly create confusion, especially in transition. What is clear, however, is that future depictions of the anatomical arrangements of the brain stem exits of CNs VI–VIII ought to reflect accurate anatomy, even though it will remain at variance with the classic numbering system. Also, Prestigiacomo [13] reported other “errors” in medical illustration and medical nomenclature exist. It is well known that CNs I and II are actually tracts, in that they do not represent the first-order sensory neurons and have a different embryological origin. Nonetheless, they continue to be referred to as “nerves.”.
The nervus terminalis, nervus intermedius, and some AA might add the motor branch of CN V, although distinct, are not numbered separately.
The Sömmering classification [10], despite many disputes during the nineteenth century, has been accepted up to the present, recently Benninger and McNeil [14] proposed a new renumeration of cranial nerves following the application of the definition of a “cranial nerve”. To be defined as cranial nerve, the nerve must originate from brainstem and pass through a foramen skull. The nucleus of the cranial nerve contains secondary sensory neurons which synapse with ganglion protoneurons or motor neurons which reach the innervated muscles. These new criteria still produced 12 pairs of cranial nerve.
Current order (Sommering) | Assessment results (Benninger and Mc Neil) | New order (Benninger and McNeil) |
| (1) Olfactory | Eliminates 1) nucleus not in brainstem, (2) primary sensory neuron | |
| (2) Optic | Eliminated, (1) nucleus not in brainstem, (2) primary sensory neuron | |
| (3) Oculomotor | Becomes1st cranial nerve | (1) Oculomotor |
| (4) Trochlear | Becomes 2nd cranial nerve | (2) Trochlear |
| (5) Trigeminal | Is split into 2 separate nerves due to separate nuclei-current sensory remains as trigeminal with ophthalmic, maxillary, and mandibular divisions as 4th cranial nerve, motor of trigeminal becomes the masticatory nerve and is now the 3rd cranial nerve | (3) Masticatory (4) Trigeminal |
| (6) Abducens | Moves to the 5th cranial nerve | (5) Abducens |
| (7) Facial | Due to separate nuclei, facial be comes 6th cranial nerve; nervous intermedius becomes the 7th cranial nerve | (6) Facial (7) Nervous Intermedius |
| (8) Vestibulocochlear | Is split into 2 nerves due to separate nuclei and separate modalities. Vestibular nerve becomes the 8th cranial nerve; and the cochlear nerve becomes the 9th cranial nerve | (8) Vestibular (9) Cochlear |
| (9) Glossopharyngeal | Becomes the 10th cranial nerve | (10) Glossopharyngeal |
| (10) Vagus | Is split into 2 divisions due to target organs: (1) Laryngopalatopharyngeal (formerly cranial root of the accessory (XI) (2) Thoracoabdominal | (11) Vagus: (a) Laryngopalatopharyngeal (b) Thoracoabdominal |
| (11) Accessory | Eliminated, nucleus not in brainstem | |
| (12) Hypoglossal | Remains the same | (12) Hypoglossal |
Table 2: Renumeration of the cranial nerves by Benninger and McNeil [14].
The cranial nerves (III-XII) according to the Sömmerring classification [14] originate in brainstem from column, 1-6, (Figure. 1) that by subdivision and migration constitute the nuclei of origin of the cranial nerves (Figure. 2).
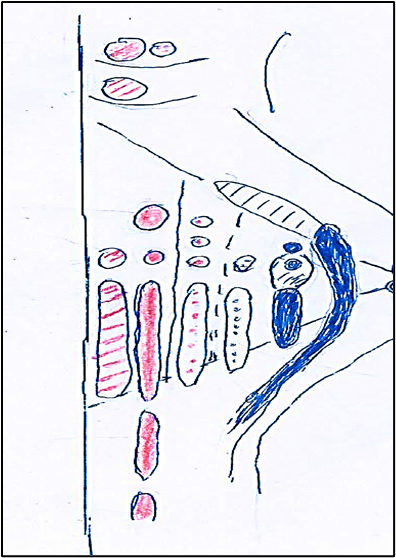
Figure 1: Columns (1-6) of the nuclei of the brainstem in medio-lateral direction. Col. 1 (red dotted lines). Column somatomotor that originates, caudo-rostrally, to somatomotor somitic nuclei: hypoglossal, abducens, trochlear, oculomotor. Col. 2 (colored in red). Column somatomotor that originates caudo-rostrally the somatomotor branchial nuclei: laryngeal, ambiguus, facial, masticatory. Col. 3 (red dots). Colomn visceromotor that originates caudo-rostrally to visceromotor nuclei: cardiopnemoenteric or ambiguus, inferior salivatory, superior salivatory, lacrimo-muconasal. Col.4 (blue dots). Column viscerosensory that, caudo-rostrally, originates the sensory nuclei: sensitive dorsal X, sensitive dorsal IX. Col. 5 (caudal blue colored; rostral dotted lines). Column viscerosensory that caudo-rostrally generates nuclei: solitary, inferior gustative, superior gustative. Col. 6 (caudal blue colored; rostral dotted lines) Column sensory that constitutesthe nucleus of the trigeminal nerve (V) which caudo-rostrally is divided into: caudal (blue) exteroceptive sensory part and rostral (dotted blue) proprioceptive sensory part.
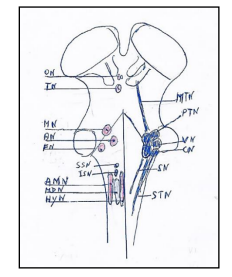
Figure 2. Brainstem nuclei with on left the motor nuclei and on right the sensory nuclei. The motor nuclei caudo-rostrally are: Hypoglossal nucleus (HyN). Motor dorsal nucleus (MDN). Ambiguus nucleus (AMN). Inferior salivatory nucleus (ISN). Superior salivatory nucleus (SSN). Facial nucleus (FN). Trochlear nucleus (TN). Oculomotor nucleus (ON). The sensory nuclei caudo-rostrally are: Spinal trigeminal nucleus (STN). Solitary nuclei (SN). Cochlear nuclei (CN). Vestibular nuclei (VN). Principal or pontine trigeminal nucleus (PTN). Mesencephalic trigeminal nucleus (MTN).
Somatotopy of Cranial Nerves: IX, X, XI, XII.
Previous our studies are regarding the somatotopy of the spinal cord [15], oculomotor complex [16], trigeminal complex [17]and facial nucleus [18], thus in this paper we report the somatotopic representation of the nuclei of the cranial nerves in the medulla oblongata: IX, X, XI, XII.
Glossopharyngeal Nerve and Nucleus
The glossopharyngeal nerve, IX cranial nerve, [19] is a “mixed nerve” with both motor and sensory fibers.
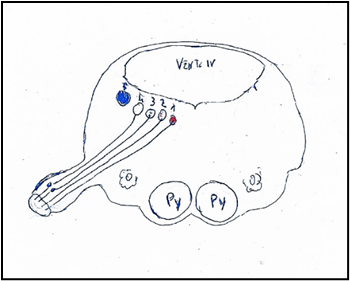
Figure 3: Nuclei of origin of the glossopharyngeal nerve mediolaterally. 1: Nucleus ambiguus. 2: Nucleus salivatory inferior. 3: Sensitive dorsal of glossopharyngeal nerve. 4: Solitary nuclei. Inferior gustative nucleus. 5: Sensory nucleus of trigeminal nerve. Py: Pyramid. O: Olive. VENT. IV: Ventricle IV.
The sensory fibers sensory convey information from the tympanic cavity, pharyngeal tympanic tube, fauces, palatine tonsils, rhino-pharynx, soft palate and the gustative fibers from 1/3 posterior or retrosulcal part of the tongue. The sensory fibers originate from the pseudounipolar neurons located in two ganglia outside the brainstem: superior ganglion or ganglion of Ehrenritter containing protoneurons which receive somatosensitive fibers or peripheral processes from auricle and send fibers to spinal trigeminal nucleus (tongue, pharynx); the inferior or petrosus ganglion or ganglion of Andersch containing the neurons: 1. neurons which receive viscerosensitive afferent fibers from fauces, carotid sinus, carotid glomus, tube and send fibers to the dorsal sensitive nucleus; 2. neurons which receive afferent taste fibers from the retrosulcal part of the tongue and project the fibers to rostral part of the nucleus of the solitary tract which is located dorsolaterally to the dorsal motor of vagus and anteriorly to the vestibular nuclei.
The motor fibers [20] originate from the cell bodies of the neurons located in the nucleus ambiguus which is situated beneath the floor of the IV ventricle, at level of the inferior olivary nucleus. The nucleus ambiguus is subdivided in the subnuclei with muscolotopic representation, so the laryngeal muscles are in caudal part of nucleus, the pharyngeal nuclei in the intermediate part, stylopharyngeus muscle and superior constrictor pharyngeal muscle in the cranial part. The glossopharyngeal nerve supplies the levator veli palatini muscle related the respiratory system, in particular with inspiration in rats [21] Recently, it was showed that the human pharyngeal muscles are heterogeneous in muscle fiber content [22] and that these fibers are arranged into two distinct and functional layers: a slow inner layer (SIL) and a fast outer layer (FOL) and that the glossopharyngeal (IX) nerveinnervates the slow inner layer, while the vagus nerve (X) innervates the fast outer layer. The two layers of fibers can be distinguished only in two-year old man, but they not found in the human newborns and nonhuman primates (monkey).
The efferent secretory parasympathetic fibers of IX nerve arise from the salivatory inferior nucleus, which is situated laterally and medially to rostral end of the ambiguus nucleus, between the hypoglossal nucleus medially and the nucleus of solitary tract laterally; these preganglionic parasympathetic fibers, by the own ramus, tympanic nerve, tympanic plexus and lesser superficial petrosal nerve arrive to “otic ganglion”. The “otic ganglion” gives origin to the postganglionic fiber which are associated with the “auriculotemporal nerve”, a trigeminal ramus, and terminate in the salivary parotid gland.
The visceral afferents of IX nerve arrive to solitary nucleus, together with the VII and X nerves.
Glossopharyngeal nerve has a communicating branch to the vagus nerve (CN X). Lesions to the CN IX are often accompanied by lesions to the CN X.
Caudal part of nucleus ambiguus gives rise to the cranial part of the spinal accessory nerve, while rostral part of this column gives rise to glossopharyngeal special visceral efferent fibers innervating the stylopharyngeus muscle.
The nerve vagus, X cranial nerve, [23] is mixed nerve containing both sensory or afferent and motor or efferent fibers innervating the larynx, pharynx and viscera (Figure. 4).
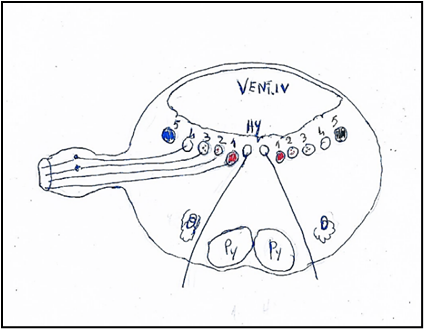
Figure 4: Nuclei of origin of the vagus nerve mediolaterally. 1. Nucleus ambiguus or Ventral nucleus of vagus nerve. 2. Nucleus cardiopneumoenteric or Dorsal nucleus of vagus nerve. 3. Sensory dorsal of vagus nerve. 4. Solitary nuclei. 5. Sensory nucleus of trigeminal nerve.
Hy: Hypoglossal nucleus. Py: Pyramid. O: Olive. VENT. IV: Ventricle IV.
The visceral afferent[24] fibers have the cell bodies located in the inferior or nodosus ganglion of vagal nerve, outside the brainstem, and arrive to the solitary nucleus and in the superior or jugular ganglion, outside the brainstem, and arrive to the trigeminal spinal nucleus.
The efferent fibers have own cell bodies in the nucleus ambiguus and the dorsal motor nucleus of the vagal nerve, both in the medulla oblongata.
The vagal fibers both afferent and efferent, are extensively distributed to the larynx, pharynx, trachea, lungs, heart, gastrointestinal tract (except the lower large intestine) pancreas, gallbladder, and liver.
The parasympathetic (dorsal motor nucleus) vagal nucleus is subdivided into nine subnuclei gathered in three regional zones: cranial, intermediary and caudal. The data in animalia suggest that the heart and lungs are represented in caudal sectors, the stomach and pancreas in the intermediate sectors, and restant abdominal viscera in the cranial and medial sectors.
The upper cervical esophagus is exerted on swallowing and peristalsis by somatic and visceral motoneurons, whereas the lower esophagus is exerted on only peristalsis by visceral motoneurons.
The results in rat [25] indicated that the upper cervical esophagus is innervated by the visceral medullary vagal motoneurons as well as the somatic spinal accessory motoneurons. The lower esophagus is innervated only by the visceral medullary vagal motoneurons.
The nucleus ambiguus (Fig 5) is the motor nucleus of both vagus and glossopharyngeal nerves. The nucleus ambiguus is located in the reticular formation about half way between the spinal trigeminal nucleus and inferior olivary complex, extends from the level of the decussation of medial lemniscus to levels through the rostral third of the inferior olivary complex; the fibers from nucleus arch dorsally, join efferent fibers from the dorsal motor nucleus of vagus nerve and emerge from lateral surface of medulla dorsal to the inferior olivary complex.
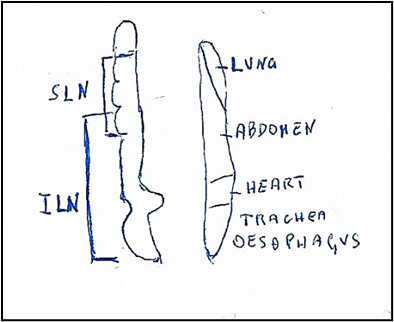
Figure 5: Nucleus ambiguus (On left). Somatotopic origin of laryngeal nerves. SLN: Superior laryngeal nerve innervating: upper oesophagyus, cricothyroid muscle, upper, middle, lower pharynx and soft palate. ILN: Inferior laryngeal nerve innervating the muscles: lateral cricoarythenoid, thyroarythenoid, interarythenoid and posterior crioarythenoid. Motot dorsal nucleus (On right). Viscerotopic localization of nuclei innervating: lung, abdomen, heart, trachea, oesophagus.
Caudal part of nucleus ambiguus gives rise to the cranial part of the spinal accessory nerve, while rostral part of this column gives rise to glossopharyngeal special visceral efferent fibers innervating the stylopharyngeus muscle.
The nucleus ambiguus in the rat [26] is made up of two major longitudinal divisions: a dorsal division, special visceral efferent component, comprising three rostrocaudally aligned subdivisions, and a ventral division, a general visceral efferent component, comprising at least two.
The dorsal division corresponds to the nucleus ambiguus in the narrow sense and comprises: a rostral compact formation part innervating the oesophagus, an intermediate semicompact formation innervating the pharyngeal and laryngeal muscles, and a caudal loose formation innervating the laryngeal muscles. Each of these formations displays a characteristic dendritic architecture. The stylopharyngeal and cricothyroid motoneurons are displaced rostral from the main pharyngeal and laryngeal motoneuronal pools. Thyropharyngeal (lower constrictor) motoneurons occupy the rostral half of the semi-compact formation and hypopharyngeal (middle constrictor) motoneurons its entire length.
The ventral division of the nucleus ambiguus corresponds to the external formation, extends along the entire length of the medulla oblongata, and contains preganglionic neurons innervating the heart and supradiaphragmatic structures innervated by the glossopharyngeal and the superior laryngeal nerves.
In the monkey [27] the nucleus ambiguus presents a column of neurons innervating the cricothyroid muscle extended from a level near the rostral end of the inferior olivary nucleus (IO) to a level caudal to its middle part. The cell columns of the cricoarytenoid, thyroarytenoid, lateral cricoarytenoid and inter-arytenoid muscles were located between the level rostral to the middle of the inferior olivary nucleus and that of the caudal end of the inferior olivary nucleus.
The nucleus ambiguus in a region of the nucleus located rostrally to the obex contains both populations overlap the motoneurons of the superior laryngeal nerve which innervates the cricothyroid muscle situated rostrally to the motoneurons of the inferior or recurrent laryngeal nerve which innervates other laryngeal muscles.
Recent studies of Pascual-Font and coll [28]. by neuroanatomical tracing study, shown in rats a more complex anatomical and functional organization; in the rat the recurrent laryngeal nerve does not contain any afferent axons from the larynx, in contrast to the pattern observed in many other species including man. The recurrent laryngeal nerve supplied only special visceromotor innervation to the intrinsic muscles of the larynx from motoneurons in the nucleus ambiguous. The superior laryngeal nerve contains afferent fibers which originate from larynx and reach the nucleus of the solitary tract.
The superior laryngeal nerve (SLN) also contained secretory efferent fibers originating from motoneurons in the dorsal motor nucleus of the vagal nerve, and special visceral efferent fibers originating from the ambiguus nucleus. Thus, this study shows that in the rat the innervation of the larynx differs in significant ways from that described in other species.
The accessory nerve [19] XI cranial nerve, emerges from the jugular foramen with glossopharyngeal and vagal nerves, but it does so by first ascending into the skull via the foramen magnum, and then exiting the central nervous system via the jugular foramen. Galen is the first to identify the accessory nerve including it with the vagal and glossopharyngeal nerves as his sixth pair. Willis [7], includes the hypoglossal nerve that is the IX cranial nerve and caudally includes the accessory nerve that is the X pair nerve referring to C1. Willis [7] excludes the accessory nerve from the encephalic nerves that is an irregular nerve because it originates entirely within the spinal cord. The Soemmerring classification [10] maintains the accessory nerve as cranial nerve XI without make sense that its nucleus and nerve begin more caudally than those of the hypoglossal or XII nerve. Benninger and McNeil [14] report that firstly Fredrici Arnold (1838) adds a cranial/bulbar root to accessory nerve and published a series of elaborately drawn anatomical plates depicting two components to the accessory nerve so that successively Henry Gray [29] in his first edition of Gray’s Anatomy reports two components of accessory nerve.
Recently Benninger and McNeil [14] believe that the structure known as the cranial root of the accessory nerve should not be included as part of the accessory nerve and it should be renamed. Furthermore, Benninger and McNeil [14] by applying contemporary embryological and anatomical findings, group the efferent peripheral nerves innervating the striated musculature into three groups: spinal, cranial and transitional. The efferent spinal nerves have a nucleus of origin within thespinal cord and innervate the musculature derived entirely from both somites and connective tissue that originates from mesoderm. The cranial nerves have nucleus in the brainstem and the same AA and also Galen, Willis and other AA propose a subdivision of efferent cranial nerves in three subcategories: 1) cranial somatic efferent (CSE) with target musculature derived from pre-otic somites (CSEpr), (oculomotor (III), trochlear (IV), and abducens (VI)); 2) cranial somatic efferent with target musculature derived from postotic somites (CSEpo) (vagus (X), Laryngopalato-pharyngeal motor (XI), and hypoglossal (XII)); 3) cranial branchial efferent (CBE) (trigeminal (V), facial (VII), glossopharyngeal (IX)), which have targeted musculature arising from somitomeres (non-somite paraxial mesoderm).
Benninger and McNeil (14) add to the categories of cranial and spinal nerves a third classification of peripheral nerve, a transitional somatic efferent (TSE) nerve, which represents the “accessory nerve proper” and combines characteristics of both cranial and spinal nerves (Table 1). Furthermore, same AA demonstrate Willis’s reasoning was correct affirming that the accessory nerve is “not an encephalic nerve” and should be regarded as a unique peripheral nerve.
Currently, the accessory nerve [30] is a only motor nerve (Fig 6) and comprises two components or parts: 1) a true cranial component, the true cranial nerve, which emerges from the medulla oblongata as a number of rootlets in series caudally with those of the vagal nerve; 2) a spinal component which arises as multiple rootlets from the anterior horn cells of C1-C5 spinal cord segments. These rootlets unite to form a single trunk which runs up within the vertebral canal and through the foramen magnum travels upward into the posterior cranial fossa and then leaves the skull through the jugular foramen and; just below the jugular foramen, the two components, cranial and spinal, separate.
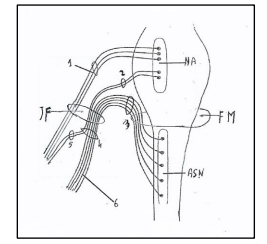
Figure 6. Nucleus ambiguus and nuclei of accessory nerve. NA: Nucleus ambiguus. ASN: Accessory spinal nucleus. 1: Nervus vagus. 2: Vagal root of accessory nerve; 3: Spinal root of accessory nerve. 4: Trunk of accessory nerve. 5: Internal branch of accessory nerve or vagal communicating branch to laryngeal muscles. 6: External branch of accessory nerve or spinal accessory nerve to trapezius and sternocleidomastoid muscles. JF: Jugular Foramen. FM: Foramen magnum. Figure modified from: Kierman J.A. Barr: Il sistema nervoso dell’uomo. A cura di Cocco L, Gaudio E, Manzoli L, Papa M. Edises p. 149. 2013.
The cranial component of the accessory nerve joins the vagus at the inferior vagal ganglion (a short distance below the jugular foramen) and thereafter ceases to have a separate identity. The cranial accessory fibers being entirely motor are distributed to the pharyngeal constrictor muscles via the pharyngeal branch of the vagus nerve.
The spinal component of the accessory nerve, SNA, runs postero-inferiorly to supply in the medial surface of the sternocleidomastoid muscle, gives ramifications to this muscle, crosses the posterior cervical triangle of the neck and enters in the inferior portion of the trapezius muscle.
The spinal component of the accessory nerve (Fig. 7) [31] comprises the group of the motoneurons supplying the sternocleidomastoid and trapezius muscles, the two occipital muscles which derivate from the lateral plate mesoderm, unlike the masticatory and facial muscles which derivate from both the cranial paraxial mesoderm and lateral plate mesoderm [32]. In rat the spinal accessory nucleus [31] longitudinally was found to be located in the caudal part (caudal 0.9-1.2 mm) of the
medulla oblongata, the whole lengths of cervical spinal cord segments C1-C5 and rostral fourth of C6. C1-C4) ([24]. In the caudal part of the medulla oblongata, the SNA was represented by a group of perikarya of motoneurons lying immediately ventrolateral to the pyramidal fibers that were passing dorsolaterally after their decussation. In the spinal cord, the motoneurons of the SNA were located in the dorsomedial and central columns at C1, in the dorsomedial, central and ventrolateral columns at C2 and in the ventrolateral column only at C3, C4, C5 and rostral quarter of C6. The perikarya of motoneurons supplying the sternocleidomastoid were located in the caudal part (caudal 0.9-1.2 mm) of the medulla oblongata ventrolateral to the pyramidal fibers that were passing dorsolaterally after their decussation. They were also located in the dorsomedial and central columns at C1, in the dorsomedial, central and ventrolateral columns at C2 and only in the ventrolateral column at the rostral three-quarters of C3. The perikarya of motoneurons supplying the trapezius muscle were located in the ventrolateral column only in the caudal three-quarters of C2, the whole lengths of C3, C4 and C5, and in the rostral quarter of C6.
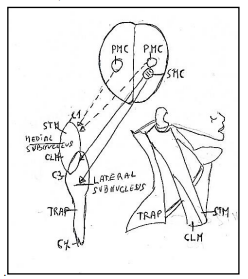
Figure 7: Subnuclei of the accessory nerve. Medial subnucleus extend between C1-C3 spinal segments innervating the muscles: sternomastoid muscle (STM) and Cleidomastoid CLM). Lateral subnucleus innervating the muscles: Cleidomastoid (CLM) and Trapezius (TRP). C1: First spinal segment. C3: Third spinal segment C7: Seventh spinal segment. PMC: Primary motor cortex. SMC: Supplementary motor cortex. Figure modified from Benninger and Mc Neil [14].
Besides, it is considered that the muscles of neck, trapezius and sternocleidomastoid, are transitional muscles [14] originated from mesoderm-derived striated muscle with connective tissue and osseous attachments that are neural crest born. These muscles are innervated by “transitional nerve” [14] having both cranial and spinal characteristics, but ultimately residing in the cervical spinal cord. Thus, the accessory nerve and its associated musculature should be regarded as a transitional nerve and transitional muscles, a new category of peripheral nerve and musculature.
The hypoglossal nerve, XII cranial nerve, [27] is a main motor nerve (Figure 8) innervating the somatic skeletal musculature of the tongue, but containing also afferent fibers.
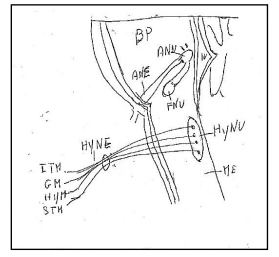
Figure 8: Hypoglossal nucleus and hypoglossal nerve.BP: Basilar Pons. ME: Medulla oblongata. IV: Ventricle IV. ANU: Abducens Nucleus. ANE: Abducens Nerve. FNU: Facial Nucleus. HyNU: Hypoglossal Nucleus. HyNE: Hypoglossal Nerve.. ITM: Intrinsic Tongue Muscles. GM: Genioglossus Muscle. HYM: Hypoglossus Muscle. STM: Styloglossus Muscle.
In cat [34] the primary afferent neurons or protoneurons of the hypoglossal nerve were located in the trigeminal ganglion, the superior ganglion of the glossopharyngeal and vagal nerves, and the first three cervical ganglia. The central projections of these protoneurons hypoglossal afferents were organized in a selective manner according to their cell origin: central projections of neurons in the trigeminal ganglion terminated in the subnuclei of the trigeminal nucleus: subnucleus dorsalis (Vpd) of the principal nucleus (Vp), lateral margin of the caudal pars interpolaris (Vi), interstitial nucleus and laminae I and V of the pars caudalis (Vc); 2. central projections of neurons in the superior ganglion of glossopharyngeal and vagal nerves terminated ipsilaterally in the caudal half of the solitary nucleus and bilaterally in the commissural nucleus [34] ;3. central projections of proprioceptive neurons in the C1-C3, first three cervical ganglia terminated in laminae I and V of the corresponding cervical cord segments [35].
In dog Chibuzo and Cummnings [36] found, by these intramuscular injections in the isolated extrinsic lingual muscles, that the primary neurons of the afferent fibers to lingual muscles, the genioglossus, hyoglossus, and styloglossus are located in the C1 spinal ganglion, proximal vagal (jugular) ganglion and trigeminal ganglion, ipsilaterally.
In monkey Kubota and coll. [37] found that the protoneurons situated in the C2, C3, C4 ganglia have the peripheral processes which, through a direct anastomosis “ansa hypoglossi or ansa cervicalis” between the cervical nerve I and the hypoglossal nerve, conduct the lingual proprioceptive impulses from the spindle muscle in tongue.
The hypoglossal nucleus originates the efferent fibers of the hypoglossal nerve. The hypoglossal nucleus forms a column of multipolar cells, about 18 mm long, that occupies the central gray of the medial eminence; it begins caudal to inferior olive and extends rostrally to region of the striae medullares of ventriculi quarti. Within the nucleus run the myelinated fibers which are the root fibers of the motor cells, the root fibers gather on the ventral surface of the nucleus, forming a series of rootlets [10-16] which pass ventrally, lateral to the medial lemniscus, and emerge on the surface of the medulla oblongata between the pyramid and inferior olivary complex.
The hypoglossal nucleus [20] is constituted of two nuclei posterior and anterior columns, each subdivided in medial and lateral subnuclei. The motoneurons have a myotopic organization, in particular the posterior subnuclei/posterolateral innervate the retrusor muscles (temporalis muscle) of the tongue; the subnuclei anterior/anteromedial innervate the protrusor muscles (medial or internal and lateral or external pterygoid muscles) of the tongue. Experimental data suggest that motoneurons of medial compartment innervate the transverse muscle (intrinsic muscles: transverse and vertical muscles and genioglossus muscle), while the motoneurons of the lateral compartment innervate the longitudinal muscles (styloglossus, hyoglossus, superior and inferior longitudinal muscles). (In proximity to hypoglossal nucleus are minor neuronal groups (perihypoglossal nuclei) that comprend the intercalatus nucleus (Staderini) with gustative and visceral connections, paramedian dorsal nucleus (reticular nucleus and the prepositus nucleus which is connected with the cerebellum and oculomotor nuclei.
In rat, the hypoglossal nucleus is subdivided in the dorsal and ventral parts. The dorsal hypoglossal nucleus [38] innervates the retrusor muscles and ulteriorly is subdivided in: a rostrolateral part with the neurons innervating the extrinsic, styloglossus muscle and a caudolateral part innervating the extrinsic hyoglossus muscle; the central and medial parts with the neurons innervating the intrinsic muscles, superior and inferior longitudinal muscles. The ventral hypoglossal nucleus [39] innervates the genioglossus muscle; this muscle in tongue of mammals has at least two subdivisions, one is horizontal and the other fans out obliquely; in dog, the hypoglossal nerve appears to have separate branches for each muscle subdivision. In rat the two subnuclei lateral and centrolateral of the ventral hypoglossal nucleus supply the two components of the genioglossus muscle.
In rat Mameli and coll [40]. showed that axons of neurons of hypoglossal nucleus also innervate the ipsilateral masseter muscle because they also spread into the masseter branch of the trigeminal nerve to target the polar portions of the masseter muscle spindles. Besides the retrograde double labelling, performed by injecting Dil into the pad and True Blue into the ipsilateral masseter muscle, showed labelled hypoglossal neurons in the medio-dorsal portion of the XII nucleus. The majority of these neurons were small (15-20micron diameter), showed fluorescence for Dil and projected to the mystacial pad. Other medium-size neurons (25micron diameter) were instead labelled with True Blue and projected to the masseter muscle. Finally, in the same area, other small hypoglossal neurons showed double labelling and projected to both structures.
A central mechanism [41] responsible for motor and premotor control of hypoglossal (XII) activity during swallowing activity, consists of the medullary and pontine populations of XII and trigeminal premotor neurons; in particular the Kolliker-Fuse nucleus has a prominent role in control of the inspiratory-related activity of the XII motoneurons supplying tongue protrusor and retrusor muscles.
In monkey macaca mulatta Morecraft and coll [42] evidenced that the hypoglossal nucleus receives inputs from extensive regions of cerebral cortex, specially from frontal, parietal cingulate and insula cortices. The hypoglossal nucleus received bilateral input from the face/head region of the primary (M1), ventrolateral pre- (LPMCv), supplementary (M2), rostral cingulate (M3), and caudal cingulate (M4) motor cortices. Additional bilateral corticohypoglossal projections were found from the dorsolateral premotor cortex (LPMCd), ventrolateral proisocortical motor area (ProM), ventrolateral primary somatosensory cortex (S1), rostral insula, and pregenual region of the anterior cingulate gyrus (areas 24/32). Dense terminal projections arose from the ventral region of M1, and moderate projections from LPMCv and rostral part of M2, with considerably fewer hypoglossal projections
Mameli and coll [43] found that the natural olfactory stimulation with amyl acetate significantly modulates the electrical activity of hypoglossal neurons and the electromyographic responses of the tongue musculature and provided the initial demonstration that olfactory information is conveyed from the olfactory bulb to the hypoglossal nucleus via the interpeduncular nucleus (IPn) by both fast disynaptic and different polysynaptic pathways. The latter, in particular, involve many of the brain structures that process olfactory information.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
