
Review Article | DOI: https://doi.org/10.31579/2690-4861/024
Department of Anesthesia, Critical Care, and Pain Medicine, Massachusetts General Hospital, Boston, Massachusetts, USA.
*Corresponding Author: Melanie M. Stipp, Department of Anesthesia, Critical Care, and Pain Medicine, Massachusetts General Hospital, Boston, Massachusetts, USA.
Citation: Melanie M. Stipp. (2020) SARS-CoV-2: Micronutrient Optimization in Supporting Host Immunocompetence. International Journal of Clinical Case Reports and Reviews. 2(2); DOI:10.31579/2690-4861/024
Copyright: © 2020 Melanie M. Stipp, This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
Received: 16 April 2020 | Accepted: 27 April 2020 | Published: 05 May 2020
Keywords: COVID-19; immunocompetence; micronutrients; SARS-CoV-2; coronavirus
The novel 2019 coronavirus (SARS-CoV-2) has become an international pandemic. The lack of targeted treatment for SARS-CoV-2 has necessitated the need to evaluate all potential modalities to enhance host immune response. This article reviews the role essential micronutrients play in the risk reduction and treatment of COVID-19. While literature supporting the risk and severity reduction of COVID-19 infections through exogenous supplementation is limited, micronutrient optimization is essential in maintaining host immunocompetence, and deficits may critically impair host defense. The immune supportive mechanisms and physiologic effects of various micronutrients are multifaceted, including the reduction of viral replication, inhibition of polymerase function, augmentation of innate and acquired immune responses, enhancement of anti-inflammatory immune responses, and the reduction of pro-inflammatory responses.
Micronutrient optimization is critical in maintaining host immunocompetence [4]. Micronutrient deficiencies may predispose the host to viral infection acquisition [4]. Severe cases of SARS-CoV-2 can trigger an uncontrolled inflammatory immune response characterized by lymphopenia, reduced natural killer cells, CD4+ cells, CD8+ cells, T lymphocytes, B lymphocytes, basophils, monocytes, and eosinophils [68]. SARS-CoV-2 induced cytokine storm triggers the excessive production of pro-inflammatory cytokines [68].Clinical laboratory findings include lymphopenia, elevated CRP, elevated LDH, elevated D-dimer and increased ferritin levels [68].Cytokine storm precipitates neutrophil and macrophage sequestration, infiltration, pulmonary hyaline membrane formation and diffuse alveolar wall thickening [68]. Subsequently, multiple organ dysfunction may ensue [68].
The immune system response consists of innate and adaptive immunity [4]. Innate immunity includes physiological barrier mechanisms of the respiratory and GI tract, and general non-specific immune responses such as inflammation [4]. This includes leukocyte activity (phagocytes, macrophages, mast cells, neutrophils, eosinophils, natural killer cells and dendritic cells) and complement system activity (opsonisation, cell lysis, agglutination, chemotaxis)[4].
Adaptive, acquired immune responses are antigen-specific, consisting of B and T lymphocyte responses [4]. B lymphocytes contribute to the humoral immune response, are highly specific, and bind directly to antigens [4]. T lymphocytes contribute to the cellular-mediated immune response through the expression of T cell receptors, and CD4+ or CD8+ cells, that recognize antigens bound to class I or II major histocompatibility complexes (MHC) [4].
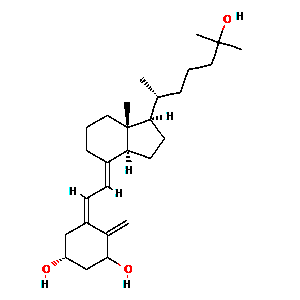
Vitamin D (1,25-dihydroxyvitamin D3)
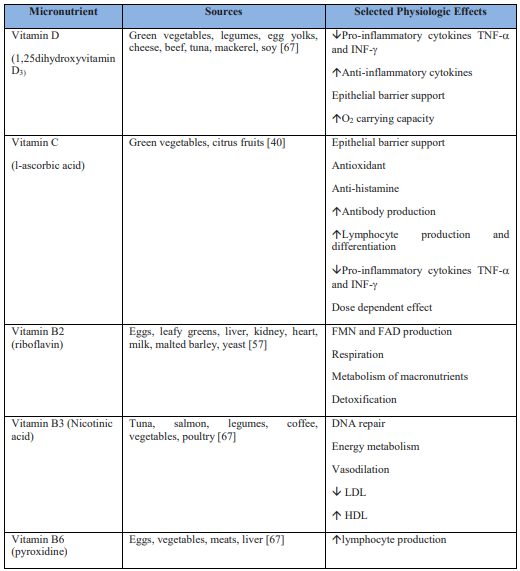
Vitamin D (1,25-dihydroxyvitamin D3) reduces the risk of host viral respiratory infection acquisition [1,5-13]. 1,25-dihydroxyvitamin D3 can be acquired exogenously or endogenously through the synthesis of steroid hormone, cholecalciferol, when the host is exposed to ultraviolet light [36, 44, 47]. Physiologically, vitamin D3 undergoes hepatic conversion to 25(OH)Dwith subsequent renal conversion to its active form, 1,25-dihydroxyvitamin D3 (1,25(OH)2D) [1].This hormonal metabolite, also known as calcitrol, directly targets genome sequences, inducing genetic and epigenetic transcription modifications [1].
1,25-dihydroxyvitamin D3 functions to promote bone mineralization by regulating serum levels of phosphorus and calcium [36]. Increased levels of serum 1,25-dihydroxyvitamin D3 provides enhanced innate immunity barrier support through the maintenance of gap, tight and adherens junctions [13]. 1,25-dihydroxyvitamin D3 reduces pro-inflammatory cytokines TNF-a and INF-g and increases anti-inflammatory cytokine expression [12, 14, 15]. 1,25-dihydroxyvitamin D3 elicits antimicrobial peptides, cathelicidins and defensins, which are found to reduce the rate of viral replication and pro-inflammatory cytokine concentrations [1,21]. Observational studies suggest a serum 1,25-dihydroxyvitamin D3 therapeutic goal of ³ 40–60 ng/mL (100–150 nmol/L) in the reduction of hospital-acquired viral infections [16-18].
Supra-therapeutic dosages of 1,25-dihydroxyvitamin D3 have been shown to reduce the duration of hospitalization in ventilated critical care patients from a hospital stay of 36 (SD=19) to a duration of 18 (SD=11) days in patients receiving treatments of 500,000 IU 1,25-dihydroxyvitamin D3 [19]. Moreover, 1,25-dihydroxyvitamin D3 has been clinically shown to increase levels of hemoglobin, enhance the metabolism of iron, and improve host oxygen carrying capacity [19-20].
Vitamin C (l-ascorbic acid)
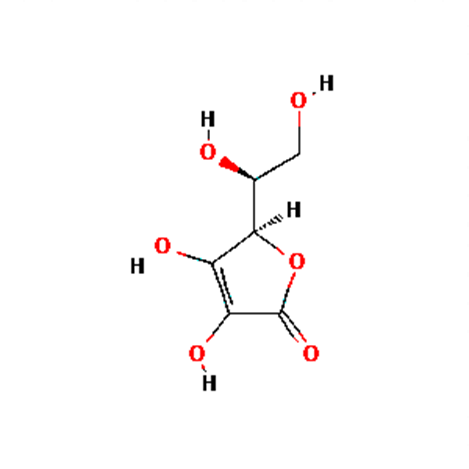
Ascorbic acid, a water-soluble form of vitamin C, functions as a potent antioxidant and detoxifier [40]. Pro-inflammatory cytokines contribute to the severe inflammation of the lungs as observed in severe cases of SARS-CoV-2 [1, 3]. Scavengers of reactive oxygen species, such as vitamin C, may play an influential role in minimizing cytokine storm and alleviating tissue damage [3, 25]. Antioxidants found in vitamin C enhance T lymphocyte response, natural killer cell activity, and interleukin-2 production [28, 29]. Vitamin C supports the maintenance of gap, tight and adherens junctions in epithelial barriers, increases serum antibody levels, and increases lymphocyte production and differentiation [22, 23]. Vitamin C has also been shown to have anti-histamine properties [44, 52].
Vitamin C deficiency has been linked to the development of viral and bacterial pneumonia [24, 32, 53]. Two randomized controlled trials identified a dose-dependent relationship between levels of vitamin C and the duration of symptomatic respiratory infections with vitamin C doses of 6-8g/day [24].
Vitamin E (α-tocopherol)
Vitamin E is a fat-soluble vitamin and antioxidant bioavailable in the form of α-tocopherol [39]. α-tocopherol is a scavenger of reactive oxygen species, neutralizing free radicals, and influential in minimizing cytokine storm and tissue damage [3, 25]. α-tocopherol prevents protein oxidation, lipid perioxidation, and inhibits protein kinase C (PKC) mediated pathways [39]. Antioxidants found in α-tocopherol enhance T lymphocyte response, natural killer cell activity and interleukin-2 production [28, 29]. α-tocopherol inhibits platelet aggregation, promotes vasodilation, stabilizes membranes, and inhibits angiogenesis [39].
Deficiency of α-tocopherol reduces adaptive immune response, hindering B and T lymphocyte function [4]. The utilization of exogenous vitamin E supplementation is controversial and may increase all-cause mortality [37]. It should be noted that simultaneous vitamin C and vitamin E supplementation has been shown to increase pneumonia and tuberculosis risk when compared to vitamin C supplementation alone [26, 27].

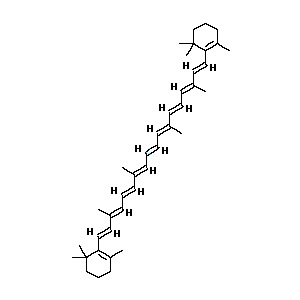
Vitamin A (β-carotene)
β-carotene is a naturally occurring retinol, pro-vitamin A [41, 46]. β-carotene has antineoplastic properties and is a scavenger of reactive oxygen species [3, 25, 41]. Antioxidants found in β-carotene also enhance T lymphocyte response, natural killer cell activity and interleukin-2 production [28, 29]. Vitamin A has been shown to reduce the overall severity and consequent fatalities associated with HIV, malaria, and measles [48]. An animal model study linked increased rates of infectious bronchitis coronavirus in chickens with vitamin A deficiency [44, 49].
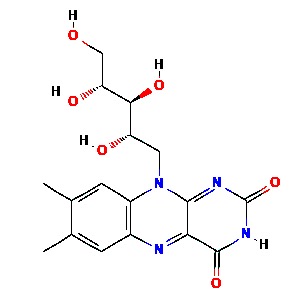
Vitamin B2 (riboflavin)
Vitamin B2 (riboflavin) is a water-soluble precursor to flavin mononucleotide (FMN) and flavin adenine dinucleotide (FAD) [57]. FMN and FAD are essential in respiration, metabolism of fats, proteins, carbohydrates, and in detoxification mediated by glutathione reductase [57]. Riboflavin and UV light have been shown to reduce human plasma viral titer levels of MERS‐CoV [44, 50].
Vitamin B3 (nicotinic acid)
Vitamin B3 (Nicotinic acid), also called nicotinamide or niacin, is a water-soluble vitamin that promotes DNA repair, energy metabolism, vasodilation, and reduced serum lipids [58]. Nicotinic acid lowers low density lipoprotein cholesterol and raises high density lipoprotein cholesterol [58]. Nicotinic acid has anti-inflammatory properties and has been shown to inhibit pulmonary neutrophil infiltration in cases of ventilator‐induced pulmonary injury, albeit, increasing overall hypoxia [44, 51]. Serum aminotransferase levels must be monitored with the administration of chronic or high dose nicotinic acid [58].
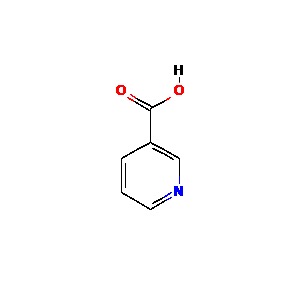
Vitamin B6 (pyridoxine)
Pyridoxine is the 4-methanol form of vitamin B6, a water soluble vitamin that acts as a coenzyme in amino acid and neurotransmitter synthesis [63]. Large dose (50-100 mg/day) pyridoxine administration has been shown to increase T lymphocyte production and activity [61, 62].

Vitamin B12 (cyanocobalamin)
Vitamin B12 (cyanocobalamin) is an immunomodulator for cellular immunity [62]. Cyanocobalamin functions to support the hematopoietic, nervous and immune systems [59, 60]. Exogenous cyanocobalamin is necessary in the myelin synthesis, cellular growth, cellular reproduction, and erythropoiesis [59, 61]. Cyanocobalamin deficiency may lead to megaloblastic anemia, disruption of neuron myelin sheath integrity, and reduced protective immune response [12]. Methyl-B12 administration has been shown to increase leukocyte, lymphocyte, CD8+ cell, and natural killer cell production (P < 0>
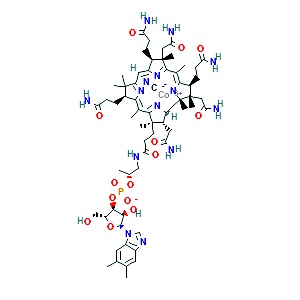
Zinc
Zinc supports both innate and acquired immune responses [44]. Zinc supplementation has been shown in several randomized controlled trials and in a meta-analysis to reduce the incidence and the duration of acute respiratory infections by 35% and 2 days, respectively. This effect has not been found to be dose dependent [30, 31]. Zinc has been shown in vitro to inhibit SARS-CoV RNA polymerase activity and viral replication [2]. Zinc lozenges are shown to reduce viral respiratory infection duration and severity if utilized within 24 hours of the onset of respiratory symptoms [30]. Zinc lozenges may reduce the binding efficacy of the SARS-CoV-2 virus to the host oral mucosa [30, 34, 35].
Iron
Iron deficiency is a contributor to the development of lower respiratory tract infections [52]. Excess iron, however, can lead to oxidative damage and viral mutation [44, 52]. Iron deficiency should be avoided in the prevention of SARS-CoV-2 acquisition. In patients with diagnosed SARS-CoV-2, elevated serum ferritin levels are observed in patients with SARS-CoV-2 induced cytokine storm [42]. One hemoglobin molecule is comprised of 2-α and 2-β subunits. Each subunit has one iron-bound heme complex [42]. Elevated serum ferritin suggests that excessive iron levels accumulate due to the SARS-CoV-2 virus interfering with the iron-bound heme complex [42]. Thus, supplementation of iron should be avoided in patients with diagnosed SARS-CoV-2 infection.
Selenium
Selenium deficiency may induce viral genome mutation of RNA viruses. These mutations may amplify virulence in the host, increasing the magnitude and severity of the resultant infection [44]. Selenium, synergistically with vitamin E, acts as a scavenger of reactive oxygen species, also influential in minimizing cytokine storm and tissue damage [3, 25, 44].
Omega‐3 Long‐chain Polyunsaturated Fatty Acids
Omega‐3 polyunsaturated fatty acids promote anti‐inflammatory effects and are precursors to protectins and resolvins [44]. Omega‐6 polyunsaturated fatty acids promote pro‐inflammatory effects and are precursors to prostaglandins and leukotrienes [44]. A delicate balance of omega-3 and omega-6 polyunsaturated fatty acids must be maintained to minimize pro‐inflammatory effects. Protectin D1 is an omega‐ 3 polyunsaturated fatty acid derived lipid mediator shown to reduce influenza viral replication, reduce influenza mortality in animal models, and has been shown to target hepatitis C viral replication [44, 55, 56].
Discussion
Anti-aging gene Sirtuin 1 (SIRT1)
Maintaining adequate micronutrient intake to optimize host immune response is indicated in both the prevention and treatment of SARS-CoV-2. Malnutrition is linked to insufficient plasma protein level of anti-aging gene Sirtuin 1 (SIRT1) [70, 71]. SARS-CoV-2 deactivates and represses SIRT1 [69]. SIRT1, dependent on nicotinamide dinucleotide (NAD+) for deacetylation, contributes to the prevention and reversal of chronic diseases that impair the host immune response, such as NAFLD, diabetes, neurodegenerative diseases and obesity [69, 70]. SIRT1 functions in global DNA reparation, fat differentiation, insulin sensitivity, neurogenesis, inflammation, genetic stability and reduces pro-inflammatory cytokines [69, 70]. SIRT1 deactivation and repression observed in the SARS-CoV-2 infection may contribute to varying levels of disease severity [69]. Micronutrient optimization in the SARS-CoV-2 infected host is necessary in the maintenance of plasma protein SIRT1 levels and in the mitigation of anti-aging gene SIRT1 inactivation [71]. Exogenous vitamin D increases plasma protein levels of SIRT1 [73]. Exogenous vitamin E has also been shown to influence plasma protein levels of SIRT1 [73]. NAD+, essential for SIRT1 deacetylation, is synthesized from its precursor, vitamin B3 (nicotinic acid) [73].
Angiotensin-converting enzyme-2 (ACE2)
Angiotensin-converting enzyme-2 (ACE2), a physiologic enzyme of the renin-angiotensin-aldosterone system (RAAS), is the receptor for the SARS-CoV-2 virus [74]. Levels of the enzyme ACE2 has been speculated as a contributor to varying degrees of the 2019 coronavirus infection severity [74]. Binding of SARS-CoV-2 virus to the ACE2 receptor results in ACE2 down-regulation, precipitating neutrophil sequestration, an exaggerated pro-inflammatory response, and a hypoxia induced release of renin [76, 77].
Vitamin D (1,25-dihydroxyvitamin D3) is a known modulator of RAAS, thus exogenous vitamin D may reduce RAAS activity through the reduction of renin expression [75, 77]. 1,25-dihydroxyvitamin D3 can be acquired exogenously or endogenously through the synthesis of steroid hormone, cholecalciferol, when the host is exposed to ultraviolet light [36, 44, 47].
Micronutrient Optimization
Vitamin D (1,25-dihydroxyvitamin D3) has been shown to reduce the risk of host viral respiratory infection acquisition [1, 5-13]. 1,25-dihydroxyvitamin D3 reduces pro-inflammatory cytokines, increases anti-inflammatory cytokines, enhances epithelial barrier support, reduces the rate of viral replication, reduces RAAS activity through the reduction of renin expression, and increases host oxygen carrying capacity [1, 13-16, 19-21]. Exogenous vitamin D should be evaluated as an adjuvant therapy in the prevention and treatment of SARS-CoV-2.
Vitamin C (l-ascorbic acid) is a potent antioxidant that may play an important role in mitigating cytokine storm and alleviating tissue damage [3, 25, 40]. L-ascorbic acid enhances epithelial barrier support, increases lymphocyte production and differentiation, and reduces pro-inflammatory cytokines [22, 23, 28, 29]. Vitamin E (α-tocopherol) is a potent antioxidant that may also play a significant role in alleviating cellular damage, enhancing T lymphocyte response, promoting natural killer cell activity, increasing interleukin-2 production, and mitigating host cytokine storm [3, 25, 28, 29]. The utilization of exogenous vitamin E supplementation is controversial and may increase all-cause mortality [37]. Concurrent exogenous vitamin C and vitamin E supplementation should also be avoided [26, 27].
Vitamin A (β-carotene) is a potent antioxidant and scavenger of reactive oxygen species shown to enhance T lymphocyte response, natural killer cell activity and interleukin-2 production [3, 25, 28, 29, 41]. Increased rates of infectious bronchitis coronavirus in chickens has been linked to vitamin A deficiency [44, 49]. Exogenous vitamins D, A, and C should be evaluated as adjuvant therapies in the prevention and treatment of SARS-CoV-2.
B vitamins also play a critical role in maintaining host immunocompetence. Vitamin B2 (riboflavin) has been shown to shown to reduce human plasma viral titer levels of MERS‐CoV [44, 50]. Vitamin B3 (Nicotinic acid) is essential for SIRT1 deacetylation [73]. Nicotinic acid has anti-inflammatory properties and has been shown to inhibit pulmonary neutrophil infiltration in cases of ventilator‐induced pulmonary injury, albeit, increasing overall hypoxia [44, 51]. Large dose (50-100 mg/day) vitamin B6 (pyridoxine) administration has been shown to increase T lymphocyte production and activity [61, 62]. Methyl-B12 administration has been shown to increase leukocyte, lymphocyte, CD8+ cell, and natural killer cell production (P < 0>
Zinc has been shown in vitro to inhibit SARS-CoV RNA polymerase activity and viral replication [2]. Zinc lozenges may reduce the binding efficacy of the SARS-CoV-2 virus to host oral mucosa [30, 34, 35]. The utilization of exogenous zinc should be evaluated for use in SARS-CoV-2 infections and initiated within 24 hours of the onset of respiratory symptoms [30].
Elevated serum ferritin in severe SARS-CoV-2 infections, suggest that excessive plasma iron levels accumulate in this state due to SARS-CoV-2 interfering with the iron-bound heme complex [42]. Exogenous iron should be avoided in these patients. Selenium, synergistically with vitamin E, acts as a scavenger of reactive oxygen species, also influential in minimizing cytokine storm and tissue damage [3, 25, 44]. A delicate balance of omega-3 and omega-6 polyunsaturated fatty acids must be maintained to minimize pro‐inflammatory effects. Protectin D1, an omega‐3 polyunsaturated fatty acid, should be evaluated as an adjuvant therapy in the reduction of cytokine induced pro-inflammatory effects and as a potential inhibitor of SARS-CoV-2 viral replication [44, 55, 56].
Conclusion
The immune supportive mechanisms and physiologic effects of various micronutrients are multifaceted. Micronutrient optimization is critical in maintaining host immunocompetence. Maintaining adequate micronutrient intake to optimize host immune response is indicated in both the prevention and treatment of SARS-CoV-2. Further investigations should be performed to evaluate the utilization of exogenous micronutrient therapies as an adjunct to the treatment of SARS-CoV-2.
Ethical Declarations
The author declares that no conflicts of interest exist.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
