
Mini-Review | DOI: https://doi.org/10.31579/2692-9406/061
1 Department of Medical Sciences, Clinical Chemistry, University Hospital of Uppsala, Uppsala University, Uppsala, Sweden.
2 Department of Cardiology, Heart Centre, and Department of Public Health and Clinical Medicine, Umeå University, Umeå, Sweden.
*Corresponding Author: Gunnar Ronquist, Department of Medical Sciences, Clinical Chemistry, University Hospital of Uppsala, Uppsala University, Uppsala, Sweden.
Citation: G Ronquist, A Waldenström. (2021) Prostasomes (Exosomes) Mediate Functional Abilities to Recipient Cells through Transfer of Proteins and Nucleic Acids: A Comprehensive Model for Exosomal Intercellular Communication. Biomedical Research and Clinical Reviews. 4(1); DOI: 10.31579/2692-9406/061
Copyright: © 2021 Gunnar Ronquist, This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 12 April 2021 | Accepted: 19 May 2021 | Published: 21 May 2021
Keywords: prostasomes; exosomes; messenger; transport; DNA; RNA prostate; heart
The prostasome is the first described exosome and constitutes the third communication system between cells mediating messages besides gap junctions and soluble compounds such as hormones. Exosomes are nanometer vesicles surrounded by a lipid bilayered membrane and released by most cell types including malignant cells. The exosomal messenger system reaches distant cells even on the other side of the blood brain barrier. In this way they are able to interact with their target cells for delivery of their cargo. We here describe prostasomal properties in more detail thus exemplifying common exosomal characteristics. Myocardial derived exosomes (cardiosomes), are also described in order to highlight other common biological functions including damaged tissue, i.e. tissue repair. Abnormal tissue such as malignant progression can be driven by cancer cell derived exosomes, believed mainly to be mediated by different forms of short RNAs exerting their action through specific signaling pathways related to metastases, therapeutic resistance and immunosuppression.
This minireview includes biogenesis, structure and some biological functions of the first described exosome, i.e. the prostasome. As many of the prostasomal characteristics are shared with those of other types of exosomes, the prostasome may serve as a symbol for other exosomes. Notably, all types of exosomes are distinct from each other dependent upon their cells of origin, i.e. the maternal cell. Some other types of exosomes will also be discussed in order to demonstrate their pluripotency.
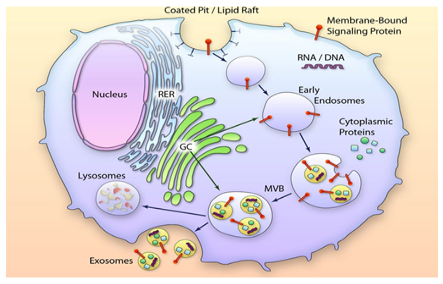
Exosomes belong to the extracellular vesicle (EV) family. They are lipid-bilayer enclosed vesicles [1] that are released from most cell types and present in all body fluids. They were first found in prostatic fluid and subsequently in seminal plasma [2-4]. They were denoted “prostasomes” [5] due to their origin in prostate epithelial cells [6, 7]. Each ejaculate contains trillions of prostasomes [8] as compared with about 150 million of sperm cells meaning an extremely high excess of prostasomes. Given their small sizes, most of them having a diameter within the range of 30-200 nm, prostasomes/exosomes are generally visualized by electron microscopy. The biogenesis of prostasomes/exosomes means multistep processes meaning two invagination events of biological membranes. The first invagination comprises the plasma membrane of the maternal cell contributing to endocytic vesicles in the formation of early endosomes that develop into late endosomes. The second invagination commences multiple inward buddings of the late endosomal membrane ending up in intraluminal vesicles (ILVs). In this way storage vesicles/multivesicular bodies (MVBs) are formed, thus retaining selected molecules from the maternal cell. The membrane surrounding the MVB can fuse with the plasma membrane of the maternal cell. Prostate epithelial cells release the ILVs as prostasomes to the ducts of the prostate gland [7, 9] (Fig 1). It should be considered that the bilayered membrane surrounding prostasomes (and all other exosomes) are ”right-side-out” with reference to the plasma membrane due to the two preceding sessions of invaginations described above. This is corroborated by e.g. Mg2+ and Ca2+ -stimulated ATPase appearing as an ectoenzyme [10] which can be verified at the outer surface of prostasomes [4-6]. Thus, the maternal cell surface interactive molecules such as enzymes and receptors do appear on the outer surface of prostasomes/exosomes.
Prostasomes as well as other exosomes demonstrate a specific lipid composition with a high cholesterol-to-phospholipid molar ratio being around 2.0 [1]. Sphingomyelin (at the expense of phosphatidyl choline) is the dominating type of phospholipid of prostasomes/exosomes constituting almost half of the total phospholipids [1, 7]. The fatty acids in sphingomyelin are mainly palmitic acid and other fatty acids consist largely of saturated and monounsaturated fatty acids [1]. For physical/chemical reasons cholesterol incorporates in biomembranes that contain saturated phospholipid acyl chains having a high content of sphingomyelin. Such a peculiar pattern suggests that the lipids in the prostasomal membrane are highly ordered. This is indeed in line with our electron spin-labeling analyses demonstrating high order parameters for prostasomes [1]. Such a highly ordered membrane structure renders prostasomes/exosomes tough withstanding physical force, e.g. freezing and thawing besides their resistance to detergents. More biologically relevant is also their ability to keep tight thus preventing leakage and protection of cargo [4].
An EV-to-target cell interaction was firstly noted between prostasomes and sperm cells [5]. This finding was followed by demonstration in free zone electrophoresis of a strong interaction between prostasomes and sperm cells exerted by hydrophobic forces [11]. The prostasomal cargo mediating effects on recipient sperm cells include various membrane proteins and nucleic acids. Among the transferable prostasomal membrane proteins is the membrane attack complex [MAC]-inhibitory protein, CD 59, also known as membrane inhibitor of reactive lysis [12]. The complement system provides the host [in this case the female genital tract] with defense against invasion of non-self components. It is activated by different pathways that all end in the formation of MAC. CD59 is expressed on the surface of many cells including human erythrocytes, but interestingly, also on the surface of an organelle, the prostasome [12, 13]. The sperm cell is continuously boosted by prostasomes with CD59, thus protecting the sperm cells in the female genital tract. The rare disease, paroxysmal nocturnal hemoglobinuria [PNH] is characterized by a lack of functional CD 59 on the erythrocyte membrane. PNH typically demonstrates an increased susceptibility of erythrocytes to complement-mediated lysis, due to absence of CD59 [12, 13]. Human prostasomal CD59 can be transferred in vitro with maintained functionality to human erythrocytes lacking CD59 (intra-species) as well as to rabbit erythrocytes also lacking functional CD59 against human complement (inter-species). Short-time incubations of such erythrocytes with human prostasomes resulted in abrogation of complement-induced hemolysis, meaning in both cases an acquired resistance to lysis and normalization of the erythrocytes through human prostasomes [13] via transfer of the CD59 glycoprotein. This demonstrates the readiness of prostasomes to deliver cargo (CD59 and Ca2+ signaling protein, see below) to recipient cells and potentially be used as therapeutic agents.
Human sperm cells are equipped by prostasomes with Ca2+ signaling mechanisms that are pivotal for the fertilization process. Freshly ejaculated sperm cells are incapable to fertilize the egg. This function is obtained only in the female reproductive tract through a functional maturation process called capacitation [14, 15]. Capacitation implicates modification in the biochemical and biophysical characteristics of sperm cells while proceeding on its way to the ovum. The process is completed when the sperm cells are recognizing ligands of the zona pellucida by undergoing the acrosome reaction [15]. Sperm Ca2+ signaling and interaction with the female reproductive tract are of utmost importance in orchestrating penetration of layers of cells and glycoproteins surrounding the egg culminating in the fusion with the egg. The detailed mechanisms of the aforementioned Ca2+ signaling have been explored by Park et al [16]. These authors observed that prostasome interaction with the sperm cell was necessary for the transfer from prostasomes of calcium ion signaling tools (e.g. receptors and enzymes) for sperm cell functionality. Sperm cells separate themselves from a majority of other cells, in that their DNA is compressed by protamine replacing histones. Sperm transcription ceases several days before the end of spermiogenesis [17]. Accordingly, expression is shut down during a time period of weeks, when sperm signaling proteins are indespensible in the female reproductive tract. As a matter of fact, sperm cells escape the hardship to produce or keep all the important signaling proteins. Simply, they get them from their “rucksacks”, (the prostasomes) on their way to the egg [16], (Fig 2). Sperm cells display a high sensitivity to progesterone, meaning that already a picomolar concentration of the hormone is sufficient for chemotactic response [18]. Park et al [16] verified that this very low concentration of progesterone was enough to induce a well-adapted, high amplitude calcium ion signal in sperm cells. Accordingly, seminal plasma with its rich supply of proteins is essential both for sperm transport and sperm protection, maturation and function [19].
DNA fragments were described in prostasomes as early as 1990 [20]. Subsequent studies revealed a chromosomal origin of the DNA, where 4 out of 13 DNA clones represented gene sequences (31%), demonstrating a staggering enrichment of exones compared to wild type cell nucleus [21]. A genome-wide DNA copy number analysis revealed that prostasomes indeed contained fragments of DNA representing the entire genome. Such DNA fragments were transferred to freshly prepared sperm cells [22]. The prostasomal/exosomal cargo mediating effects on recipient cells also include messenger and microRNAs (miRNAs) [23]. miRNAs function as regulators through degradation or inhibition of specific mRNA targets [24-26]. This functional pattern is not exclusive for prostasomes but includes other exosomes as well [27]. Furthermore, fragments of transfer RNA can in a non-specific way interfere with protein translation or function similar to miRNAs by binding components of the RNA-induced silencing complex [28]. The prostasomal content of miRNAs is substantial [8]. This is contrary to findings in exosomes isolated from blood plasma and cell culture supernatant, where the majority of extracellular miRNAs appears independent of exosomes [29]. Also, prostasomes exhibit a specific miRNA profile that differs from the profiles of exosomes of other origins. This indicates the specificity of exosomes based on the maternal cell, i.e. cell of origin [8]. The female reproductive tract is a hostile environment to sperm cells due to their “non-self” existence in that location. Prostasomes with their unique cargo may act as sperm cell protectors by their immunomodulatory ability [8]. The survival of sperm cells is in this way prolonged in an otherwise hostile environment in favor of a successful fertilization. The detailed description of prostasomal interactive function with its natural target cell (sperm cell) may serve as reference also for other exosomal interactive relations with the respective target cells.
Accordingly, the prostasome is only a part of a broader exosomal messenger system in mammals. This is demonstrated by e.g. the production of cardiosomes from cardiomyocytes. Cultured rat cardiomyocytes are able to release cardiosomes and when incubated with fibroblasts do penetrate the fibroblast plasma membrane. It was demonstrated that such cardiosomes contained both DNA fragments and RNA and that the DNA fragments corresponded to the whole genome [30]. The DNA was transported all the way to the nucleus of the fibroblast. Furthermore, the cardiosomes induced numerous changes in gene expression of the fibroblast. Thus exosomes do convey biological intercellular messages. Moreover, it was demonstrated that the external milieu of the maternal cell is decisive for the specific capability of the exosomes released. Thus, when cultivated cardiomyocytes were subjected to different growth factors the properties of the cardiosomes released differed such that the genetic expression of the recipient fibroblast induced both up and down regulations [31]. These in vitro results were later implemented in a whole animal model. It is well known that subjecting a heart to repeated short periods of ischemia with reperfusion in between renders the myocardium more resistant to subsequent longer periods of ischemia. This phenomenon is denoted ischemic preconditioning (IPC). Also, intermittent ischemia of a limb may protect the myocardium from ischemic attacks and is denoted remote preconditioning. The two preconditioning phenomena are mainly mediated by muscle exosomal interference. This implies that it is not mandatory that the influencing exosome originates from the same type of exposed muscle, thus skeletal muscle exosomes can successfully interfere with myocardium. In an in vivo model the beating heart of anesthetized pigs were subjected to IPC. Exosomes were extracted from the coronary sinus draining the heart from blood aiming at high cardiosomal yield. The findings demonstrate that IPC influences the mRNA in cardiosomes including gene transcripts coding for proteins with protective effects in IPC [32]. Preconditioned maternal cells release cardiosomes containing mRNA sequences that are different from mRNA content in cardiosomes from non-preconditioned cells. This finding demonstrates the importance of the external milieu on maternal cell exosomal release. As expected, neither exosomal content nor quality of DNA sequences change after preconditioning [33]. The seminal role of exosomes to mediate protective factors in the IPC and remote IPC phenomena is obvious.
It was shown in 1999 that prostasomes were found also in neoplastic epithelial prostate cells [34, 35, 36]. Hanahan and Weinberg [37] conceptualized cancer with hallmarks of the malignant disease in the following 6 points:
All these abilities require cell to cell communication. The traditional view on the mode of communication was gap junctions between cells and soluble growth factors. Today it is obvious that exosomes are a third communicative factor [38, 39]. Cancer cell derived exosomes are pertinent in all the 6 hallmarks above. As a matter of fact malignant progression can be driven by cancer cell derived exosomes. The component of such exosomes, as has been mentioned above, include proteins, lipids, DNA, mRNA, miRNA, long non-coding RNA and circular RNA that can transform the tumor extracellular matrix microenvironment. The tumor derived exosomes mediate in this way “functional components in order to initiate pathways that are necessary for tumor survival and propagation” [37]. Their mode of action to stimulate tumor growth and development is through specific signaling pathways related to metastases, therapeutic resistance and immune suppression. Cancer cell derived exosomes promote formation of metastases by initiating epithelial-mesenchymal-transition (EMT) within the tumor microenvironment. They will also travel to distant places (even passing the blood brain barrier) via various interstitial fluids in the extracellular space and be selectively taken up and induce transformation of normal cells into malignant cells [40-44].
Exosomes comprise a hitherto extensively underestimated messenger and transport system including the whole organism. They constitute the third communicative system for both near and distant interaction between cells besides gap junctions and soluble molecules. On top of this they also serve as a supplier system.
Exosomes represent a third type of intercellular communication- and supplier system.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
