
Research Article | DOI: https://doi.org/10.31579/2690-1897/199
NYU Grossman School of Medicine, New York, NY 10010
*Corresponding Author: Aubrey C. Galloway, MD Seymour Cohn Professor Department of Cardiothoracic Surgery NYU Grossman School of Medicine 530 1st Avenue, Suite 9V, New York, NY 10016.
Citation: Michael P. Dorsey, Navneet Narula, Harvey I. Pass, Les James, Valeria Mezzano, et al, (2024), post-COVID-19 Aortitis with Acute Thoracic Aortic Aneurysms: A Gene Expression and Molecular Pathway Analysis, J, Surgical Case Reports and Images, 7(7); DOI:10.31579/2690-1897/199
Copyright: © 2024, Aubrey C. Galloway. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 14 June 2024 | Accepted: 03 July 2024 | Published: 14 August 2024
Keywords: digital spatial profiling, covid19, inflammatory thoracic aortic aneurysms
Post-COVID-19 aortitis (AoC19) with aneurysm formation has been described, but little is known about the molecular pathophysiology. This study utilized Digital Spatial Profiling (DSP) to evaluate differentially expressed genes (DEG) and molecular pathway activation in the aorta of AoC19 compared to Giant Cell aortitis (AoGC).
Methods: Histopathology analysis was performed on AoC19 and AoGC and regions of interest (ROI) based on varying degrees of inflammation were chosen and encircled. ROIs were stained with fluorescent anti-cytokeratin, anti-leukocyte, anti-nuclear antibodies and profiling agents for 1,850 RNA targets using the COVID-19 Immune Response Atlas. DSP was performed to compare DEG in AoC19 compared to AoGC, and across zones of AoC19 itself. Molecular pathway activation and gene primary ontology functions were analyzed.
Results: 103 DGEs were demonstrated when comparing AoC19 to AoGC. In AoC19, the pathways activated were associated with viral protein interaction with cytokines and cytokine receptors, cytokine-cytokine receptor interactions, and COVID-19; while in AoGC, pathways activated were associated with human leukocyte antigens HLA-DRB1, HLA-DRB4, and CD4 T cells. 42 DGEs were demonstrated when comparing ROIs across AoC19 itself, activating molecular pathways associated with cytokine-cytokine receptor interactions, complement and coagulation cascades, signaling pathways for TNF, IL-17 and nuclear factor-kappa ß, and rheumatoid arthritis. The primary molecular ontology functions of the DGEs across AoC19 were for COVID-19, Kawasaki Disease/Takayasu arteritis, and aneurysm formation.
Conclusions: This study demonstrated differential gene expression in the aortic wall of AoC19 with the activation of molecular pathways shared between COVID-19, Kawasaki Disease/Takayasu arteritis and aneurysm formation.
AoC19 aortic aneurysm specimen from patient with post-COVID-19 aortitis
AoGC aortic aneurysm specimen from patient with Giant Cell aortitis
COVID-19 coronavirus 2019 disease
DEG differentially expressed genes
DSP digital spatial profiling
FDR false discovery rate
KEGG Kyoto Encyclopedia of Genes and Genomes
ROI regions of interest

Patients with COVID-19 can develop serious cardiovascular complications resulting in increased morbidity and mortality, including acute myocardial infarction, myocarditis, vasculitis, vascular thrombosis and thromboembolic events.[1,2] The SARS-CoV-2 viral spike (S) protein enters cells through angiotensin-converting enzyme 2 (ACE2) receptors, which are highly expressed in the heart, vascular system, respiratory tract and gastrointestinal tract. RNA sequencing data from multiple vascular beds, however, has demonstrated a low level of ACE2 expression in vascular endothelial cells compared to epithelial cells in the respiratory track and enterocytes in the gastrointestinal tract.[3-6] Therefore, much of the vascular endothelial cell damage in COVID-19 may be mediated by the host inflammatory immune response through the activation of inflammatory cytokines, macrophage and lymphocyte infiltration, and platelet and complement activation. The SARS-CoV-2 viral spike (S) protein also binds directly to toll-like receptors and nucleotide-binding and oligomerization domain (NOD)-like receptors on host leukocytes to activate the host immune system directly. In the most severely ill COVID-19 patients the host immune response was found to be dysregulated and maladaptive, which contributed to multisystem organ damage and the high mortality in these patients.[7] The host immune transcriptomic response signature to COVID-19 can persist for 6 weeks to 8 months after the initial infection and, result in late post-acute COVID-19 sequelae in certain patients.8,9 Late sequelae most commonly occurs 2-3 months after the initial infection, and the etiology is thought to be due to a persistent, maladaptive host immune response pattern rather than ongoing infection.[9] The development post COVID-19 aortic aneurysms has been reported as a rare late complication, although the molecular mechanism is unknown. [10,11,12]
This study was generated after we treated a patient who presented with large vessel aortitis and acute thoracic aortic aneurysms six weeks after a mild COVID-19 respiratory illness. Based on the temporal relationship to the initial COVID-19 infection, and the unusual operative findings of acute inflammatory thoracic aortic aneurysms, with areas localized dissection, acute thrombus, and multiple pseudoaneurysms, our hypothesis was that this likely represented a late post-acute COVID-19 sequelae of large vessel inflammatory aortitis (AoC19). This study was designed to evaluate gene expressing and the molecular pathway activating patterns in the aortic wall of this patient (AoC19) using Digital Spatial Profiling (DSP) and to compare this to patient with Giant Cell aortitis (AoGC).
Patient Population
A 59-year-old female with a history of hypertension, diabetes, obesity, and metabolic syndrome presented with mild upper respiratory symptoms, normal oxygen saturation scattered mild infiltrates on Chest X-ray, a normal mediastinum, and tested positive for COVID-19 on nasopharyngeal swab. She did not require oxygen or hospital admission and was discharged. At home she remained clinically well, without further symptom, and apparently recovered, but developed left chest pain and back pain 6 weeks later and represented to the emergency room. The follow-up chest x-ray revealed clearing of the prior pulmonary infiltrates, but demonstrated a newly enlarged mediastinum. The nasopharyngeal swab for COVID-19 was negative, while blood work demonstrated anti-COVID-19 immunoglobulins (IgG.3.4, IgG 16.9, and IgM 3.9) consistent with a prior COVID-19 infection, and elevated inflammatory markers (D-dimer 1258 ng/mL, Ferritin 1035 ng/mL, and C-reactive protein 220 mg/L). Work up for infection (blood cultures, human immunodeficiency virus) and autoimmune disease (antinuclear antibodies, rapid plasma reagin, and antiphospholipid antibodies) was enegative. A computed tomographic angiography (CTA) of the chest demonstrated diffuse thickening of the thoracic aorta consistent with aortitis, and saccular pseudo-aneurysms in both the ascending and descending thoracic aorta. (Figure 1) Due to ongoing left lower back pain she underwent urgent endovascular aortic repair (TEVAR) of the descending thoracic aorta. Her pain resolved and she was discharged home on a course of steroids for presumed inflammatory aortitis, with plans for follow-up elective open repair of the ascending aorta and aortic arch in 2-3 weeks. However, she developed recurrent chest pain in 2 weeks and was readmitted for urgent surgery for ascending aorta and zone 2 aortic arch replacement. The operative findings included extensive inflammation and thickening of the aorta, with multiple areas of localized dissection, thrombus, and pseudoaneurysms. (Figure 2, B, C) She recovered uneventfully and was discharged home on postoperative day 5. The postoperative CT scan demonstrating the ascending aorta and zone II aortic arch replacement is shown in Figure 3.
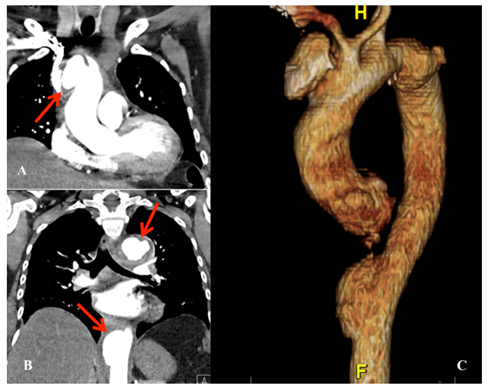
Figure 1. CTA coronal view with arrow demonstrating 4.5cm saccular pseudoaneurysm of the ascending aorta CTA coronal view with arrows demonstrating two descending thoracic aortic pseudoaneurysm (arrows) CTA three-dimensional reconstruction demonstrating multiple thoracic aortic pseudoaneurysms.
CTA, computed tomographic angiography
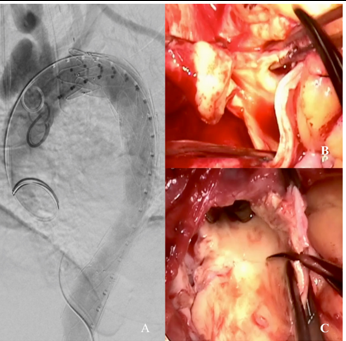
Figure 2. A. Completion aortogram after initial endovascular repair of the descending thoracic aorta. B. Intraoperative image demonstrating the whitish, inflamed and thickened ascending aorta. C. Intraoperative image demonstrating intimal thickening and multiple penetrating defects in the inner curvature of the open aortic arch.
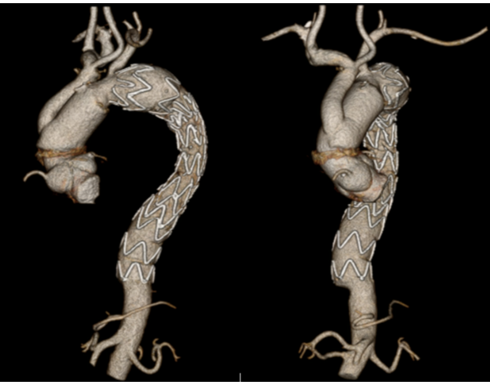
Figure 3. Postoperative CT scan demonstrating the ascending aorta and zone II aortic arch repair with debranching, and prior descending thoracic aorta stent graft. CT (computed tomographic).
This study was conducted under the NYU Grossman School of Medicine Lung Cancer Biomarker Center (#I8896) and underwent expedited IRB review and approval in 2021. Histopathologic analysis, along with special bacterial and fungal stains, was performed on the AoC19 specimen, and on a specimen from a patient with Giant Cell aortitis (AoGC) who had previously undergone ascending aneurysm repair. Electron microscopy to test for viral particles was also performed on the AoC19 specimen. Regions of interest (ROI) in different zones of the aortic, based on varying degrees of inflammation or fibrosis, and were marked and encircled for subsequent analysis. Formalin-fixed, paraffin-embedded sections of the specimens were subjected to a standard antigen retrieval protocol and stained with fluorescent anti-cytokeratin (i.e. pan-cytokeratin), anti-leukocyte (i.e. CD45) antibodies, and DNA binding SYTO-13 for nuclear recognition. The barcodes for each fluorescent DNA target in the ROIs were then selected for analysis and decoded for sequencing. (Figure 4) DSP) was then performed to evaluate DEGs, using the COVID-19 Immune Response Atlas (NanoString, WA, USA), which included a combination of the GeoMx Cancer Transcriptome Atlas of 1850 immune genes with an additional 30 genes from a specially designed COVID-19 panel. DEG analyses were performed by comparing ROI’s in AoC19 to AoGC, and by comparing ROIs across various zones across
AoC19 itself, representing normal areas, and zones with varying degrees of inflammation or fibrosis. Log fold changes with a false discovery rate (FDR) of <0>
The host inflammatory process plays a major role in the development of thoracic aortic aneurysms, producing endothelial cell dysfunction, vascular smooth muscle cell stress with a change in phenotype, and the activation of matrix metalloproteases in the extracellular matrix. (13) An analysis of bulk transcriptomome and single-cell RNA sequencing data identified 1525 DEGs in thoracic aortic aneurysms compared to normal aortic controls, activating multiple immune cell types, including macrophages, neutrophils, and T cells, and molecular pathways involved with leukocyte trans-endothelial migration, and the activation of complement and coagulation cascades. Thirty-nine (39) DEGs were identified in the macrophage population of and 30 DEGs in the T cell population. These were associated with increased infiltration of M2 macrophages and CD8+ T cells into the aortic wall, suggesting a major role of these cell types in development of thoracic aneurysms. (13)
Large vessel vasculitis with inflammatory thoracic aneurysms are most commonly due to Giant Cell aortitis (AoGC), Takayasu’s arteritis, and isolated aortitis. (14,15) While the exact etiology is unknown, the molecular pathophysiology is thought to be antigen related, with the activation of inflammatory cytokines, macrophages, and T cells (13) Using global transcriptomic profiling and bulk RNA sequencing, Hur and associates compared DEG signatures between inflammatory and non-inflammatory aneurysms, and demonstrated 159 upregulated DEGs and 93 downregulated DEGs in the inflammatory group. (16) Gene ontology enrichment demonstrated that the top 3 gene ontology terms were for immune response, host defense response, and inflammatory response; and pathways associated with increased cytokine and chemokine activity, and autoimmune disease. (16). Patients with Giant Cell aortitis have been shown to have DEGs in the aortic wall and circulating leukocytes that activate CD4 T cells and macrophages, and molecular pathways associated with a Type I interferon signature.[17]
Multisystem inflammatory syndrome in children (MIS-C) is a systemic medium vessel vasculitis thought to be triggered by SARS-CoV-2. It typically develops 3-4 weeks after the initial infection, (18,19,20) MIS-C) and is distinct from Kawasaki Disease, but with overlapping transcriptomic pathways. (18). An artificial intelligence-guided review of host immune signatures in the COVID-19 pandemic revealed shared host immune response patterns between COVID-19 (SARS-Co-V-2), MIS-C, and Kawasaki disease. (20).
The patient in this study presented 6 weeks after a mild COVID-19 infection with acute large vessel vasculitis and thoracic aortic aneurysms. There were no viral particles in the aortic specimen and the patient did not have an active infection. Our findings, however, demonstrated DEGs in the aortic wall that activated KEGG pathways associated with viral protein interaction with cytokines and cytokine receptors, cytokine-cytokine receptor interactions, and COVID-19 with primary ontology functions for COVID-19, Kawasaki Disease/Takayasu arteritis, and aneurysm formation. These findings suggest a shared and overlapping expression of host immune transcriptomic pathways in large vessel inflammatory vasculitis, inflammatory thoracic aneurysms, MIS-C, Kawasaki disease and autoimmune aortitis, The study is unique in the use of multiplexed DSP to analyze DEGs and molecular pathway activation patterns in specific areas of the aortic wall. This approach is considerably different than the more widely used bulk transcriptomic and single cell RNA methods, which measure DEGs in the entire aortic specimen. Based on this pilot study it is possible that DSP might be useful in analyzing molecular mechanisms in specific zones of aortic wall in various types of aneurysms, which could lead to a better understanding of the molecular pathophysiology involved.
This was a pilot study with only one patient and one control, and has many limitations. Based on the small sample size, the statistical significance of the findings should be considered marginal and the findings need to be validated across a larger number of patients. A major limitation is that the study does not have a normal aorta as a control. These findings only represent DEG between specific areas within the aortic wall across the AoC19 specimen, and DEGs in comparison to Giant Cell aortitis, and do not identify DEGs compared to a normal aorta. While DSP may have certain advantages in allowing the spatial comparison of gene expression in the microenvironment of tissue specimens, that is also a limitation, and the use of bulk transcriptomics and single cell RNA may be more robust in identifying major tissue wide transcriptomic patterns across a large number of specimens or patients.
Pass, Mezzano, Zhou, Tsirigos, Ramaswami, James, Peterson, and Smith, have nothing to disclose. Dr. Galloway has intellectual property with Medtronic and receives royalties for valve repair devices.
This study was supported by NYU Grossman School of Medicine, Department of Cardiothoracic Surgery Research Fund. The Experimental Pathology and Genome Technology Center core laboratories are a part of the NYU Grossman School of Medicine, Division of Advanced Research Technologies, and are funded in part by the National Institutes of Health/National Cancer Institute [5 P30CA16087].
Abstract presented at 2022 AATS Aortic Symposium: “Post-COVID-19 Aortitis with Aneurysm Formation.”
The study was conducted under the NYU Grossman School of Medicine Lung Cancer Biomarker Center (#I8896) and underwent expedited IRB review.
The GeoMx gene tag sequencing data sets generated in this study will be available at GEO, and Code will be available at GitHub. Microscopy image data is stored in public OMERO Plus and will be accessible through the NYU Data Catalog upon publication.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
