
Review Article | DOI: https://doi.org/10.31579/2690-8794/015
*Corresponding Author: Eman Refaat Youness, Department of Medical Biochemistry, National Research Centre, Egypt
Citation: Eman R Youness (2020) Overview on Epigenetics and Cancer,2(3): DOI: 10.31579/2690-8794/015
Copyright: © 2020 Eman Refaat Youness, This is an open-access article distributed under the terms of The Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 19 February 2020 | Accepted: 27 February 2020 | Published: 09 March 2020
Keywords: epigenetics,chemical processes,cancer
Epigenetics is considered as the science of hereditary phenotype which does not encompass amendment in the DNA. This occurs through chemical processes that modify the phenotype, without altering the genotype. A large number of studies showed that metabolic diseases are highly associated with epigenetic alterations suggesting that epigenetic factors may play a central role in cancer. Recent advancements in the rapidly evolving field of cancer epigenetics have shown extensive reprogramming of every component of the epigenetic machinery in cancer including DNA methylation, histone modifications, nucleosome positioning and non-coding RNAs, specifically microRNA expression. Studies of the mechanism(s) of epigenetic regulation and its reversibility have resulted in the identification of novel targets that may be useful in developing new strategies for the prevention and treatment of cancer.
Epigenetics is considered as the science of hereditary phenotype which do not encompass amendment in the DNA [15]. This occurs through chemical processes that modify the phenotype, without altering the genotype[18]. In epigenetics, the Greek acanthus epi-(over) behold countenance which is "on the top of" i.e "in addition to" the conventional basis of inheritance of genetic [49]. Epigenetics frequently betoken changes that affect gene expression and activity. Also, it is used to depict each change in the heritable phenotype. Any effect in the physiological & cellular phenotypic traits could arise by environmental and external factors [6].
This means pertinent changes to the genome but not involve an alteration in the sequence of nucleotide. Expression of genes could be well-ordered via the repressor proteinswhich bind to silence regions in DNA. These alterations possibly will remain over cell divisionsfor the period of the cell's life. It could also continue in multiple generations while they include no variations in the sequence of DNA of the organism [7].

Epigenetic alterations amend definite genes activation, not the genetic code DNA sequence. The structure of DNA or the related chromatin proteins may be adjusted, producing either silencing or activation. This mechanization allows cells to make the genes that are essential for their own activity only. Epigenetic alterations conserved during cell division. The majority of epigenetic alterations occur only within lifetime of the organism. These alterations could be patrimonial to progeny via a process termed trans-generational epigenetic inheritance. Additionally, whether gene inactivation occurs in an egg or sperm which leads to fertilization, this epigenetic modification could be transmitted to the following offspring [8].
Epigenetics and development continue from solitary impregnated embryo into immeasurable various tissues and cells [39]. It was notable that more modern and accurate epigenetic concept could be observed early in variegating eye colour in Drosophila which changes from generation to another [29]. In yeast, the effect of telomere location found like the idea of semen as it switches among phenotypes happens with no genotypic variation [19]. Epigenetics still in its infancy, by the persistent appearance of new states (bivalent promoters), novel players (lncRNAs) and novel histonemodification [39].
DNA is compacted in chromatin and nucleosome is its basic unit. Each nucleosome is formed of histone. This histone surrounded by about 145–147 DNA base pairing. Histone designed as four proteins couples, named H2A, H2B, H3, and H4. DNA is firstly “supercoiled” and then wrapped onto histones, to allow their compressing into a single cell [33].There are two types of chromatins rendering to the grade of winding of DNA. DNA in euchromatin, is extra relaxed allowing genes transcription, & DNA in heterochromatin is super-coiled and cannot accessed for gene transcription (Figure.2).
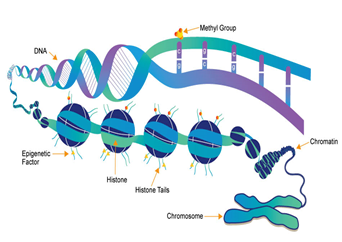
There is equilibrium between euchromatin & heterochromatin that authorize the cells to amend gene expression. This leads to notable alterations in their biological and functions phenotypes[58]. This merit achieves from the same DNA sequence; cells might discriminate in actual dissimilar means. The renovation between euchromatin and heterochromatin is called epigenetic regulation[43].Sequence; cells might discriminate in actual dissimilar means. The renovation between euchromatin and heterochromatin is called epigenetic regulation [43].
DNA methylation
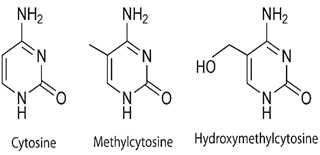
Methylation of DNA is firmly concomitant with parental imprinting, circumstantial regulation of gene expression and time, silencing of repetitive elements, inactivation of X-chromosome, human diseasesand cell differentiation (Tirado-Magallanes et al., 2017). Chemical process in the methylation of DNA, a CH3 group is linked together with cytosine residue at C5, producing 5-methylcytosine (5mC) (Figure 3).

Methylation of DNA is implement by DNA methyltranferase (DNMTs) certain sequences in DNA specially rich in guanine & cytosine nucleotide pairs, termed CpG areas, are the idealgoal of DNMTs. CpG regions in DNA of humans, (5'—C—phosphate—G—3') are not distributed at random. They are focused in small enriched CpG DNA morsels, termed CpG islands. Around 60% of all promoters in genes are wealthy of CpG islands, so these genes can be regulated epigenetically. There are 2 types of DNMT. DNMT1, which conserves predating manner of methylation next DNA replication in the cells, DNMT3A & DNMT3B, called “de novo” DNMT, that methylate earlier unmethylated DNA [43].
Demethylation of DNA could be passive, a method emerging due to DNA replication deprived of de novo methylation coupling, or active; this including the interposes 5-formylcytosine (5fC), 5-hydroxymethylcytosine (5hmC), also, 5-carboxycytosine (5caC) [47]. The active demethylation be contingent on protein complexes (demethylase) as base-excision repair (BER) and thymine DNA glycosylase (TDG) [26].
General patterns for DNA methylation that manipulate the gene expression regulation are abridged herein figure (4) [24].
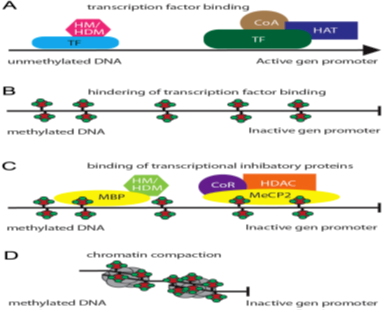
Genes that transcribed actively are generally conspicuous hypo-methylation around the transcription start point. The intricacy of the link between methylation and DNA transcription was demonstrated through genes together with promoters possessing decreased content of CpG region and hypermethylation that transcriptionally active; e.g., in gamete formation [21]. Furthermore, genes that are transcriptionally active are usually hypermethylated in the gene of the body [4]. This is supposed to chunk the start of false transcription, promote production of abnormal RNAs [40].
Human being genome comprises about fifty six million sites for CpG, about 65–75% of them be methylated [28]. During genome, sites of CpG are distributed unequally. CpG islands are demarcated as zones with minimum size, 550 b.p [12]. During the mammalian life cycle, epigenetic marks endure wide reprogramming and methylation of the CpG site, both are developmentally planned and gained environmentally. They reprogrammed in general after fertilization throughout early gametogenesis and in the zygote proximately[55]. Reprogramming includes wiping off of most DNA methylation that followed by the formation of newgroups of marks of DNA methylation [37]. The wiping off of DNA methylation throughout reprogramming was wide-ranging however, some DNA methylation continues and smooth the way for potential epigenetic inheritance [59].
DNA Hydroxymethylation
Hydroxymethylation of Cytosine was concealed as another significant epigenetic modification of DNA in mammals. Like methylation, hydroxymethyl group replaces the hydrogen atom at the cytosine position-C5, (fig 5).
It has been elucidated that hydroxymethylation of cytosine is involved also in gene regulation. e.g., the hydroxymethylation level was linked with the stem cells pluripotency. Disturbing of DNA cytosine hydroxymethylation could lead to jumbled cell functions; give rise to altered species of cancers, such as cancers of myeloid [45].
Histone and DNA
Proteins termed histones are that relate the DNA in chromosomes in eukaryotes forming basic units named nucleosomes. The DNA double helix is swathed around histones. Then these nucleosomes packed together to form chromatin. Chromatin appears as visible stainable mass under microscope during cell division. This chromatin comprises the chromosome bounded by non-histone and histone proteins [1](Fig 6).
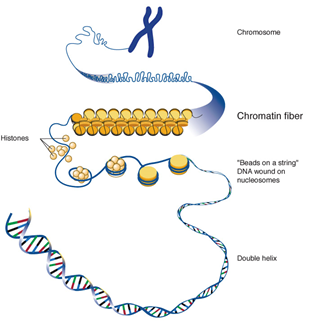
In mammals, all chromatin structure can be inherited is a form of epigenetic inheritance since it does not require the direct modification of DNA. The highly condensed form of chromatin, heterochromatin, can switch off any genes in this way. This type of chromatin can be effective as an epigenetic mechanism, as the DNA is stiffer to access to be transcribed. In the other type of chromatin, euchromatin, there is an open chromatin and the genes are ready to be accessed. It was found that genes in heterochromatin are commonly silenced and active in euchromatin [36].
Just as DNA is methylated, histones may also be modified, by methylation, phosphorylation, acetylation, ADP ribosylation and proline isomerization, but these methods of histone modification have not all presented an obvious epigenetic effect similar to DNA methylation [52].
Histone modifications
Histone covalent modifications play an imperative role in epigenetic regulation.This occurs though binding of different enzymes to histones causing phosphorylation, acetylation, methylation, or ubiquitylation to them. These modulations result in gene transcription & chromatin relaxation. On the other side, chromatin compaction & gene repression occur. Whereas DNA methylation cause silence to histone modifications, gene expression, are capable for activation or silencing of genes. This depends on adding residues on the beleaguered histones, finally on the degree of the modulation [55]. E.g., lysine acetylation at the N-terminal of histone is capable to eradicate the histones positive charge, allowing negatively charged DNA to be detached from them and gaina transcription-ready arrangement. Furthermore, methylation of histonelysine leads to transcriptional activation or suppression. This depends on the position of methylated lysine. Acetylation of histones is interposed by(HAT) histone acetyltransferase, whereas deacetylations are encouraged by (HDAC) histone deacetylases. In acetylation and methylation, the preferred targets are the H3 and H4 center histones the same as the enormous preponderance of covalent modifications include them. In general, H4K20 methylation, H3K27 and histone H3K9 (lysine in position 9 ofthe H3 histone), are related with repression of gene. Conversely, methylation of H3K36 & H3K4, also, acetylation of H4 &H3, categorizes zones of active gene expression [33].
Regulation of miRNA
Mi- RNAs (mi-RNAs) are short non-coding RNAs (18–20 nucleotides), that join messenger RNA (mRNAs) through complete or incomplete matching their 3' untranslated zones and resulting in their deprivation and post-transcriptional gene silencing. Various mi-RNAs positions are situated in the intron regions. Mi-RNAs are capable to permit epigenetic control on apoptosis, and other fateful biological processes [14].
Abnormalities of epigenetic in tumorigenesis
Mechanisms of epigenetics are in demanded for preserving normal development, growth & expression of genes in various organs. Abnormality of epigenetic regulation may change the expression of genes and functions which could result in diseases as cancer. During formation of cancer, a large number of mutated genes are produced or abnormally activated [9].
There is equilibrium between euchromatin & heterochromatin that authorize the cells to amend gene expression. This leads to notable alterations in their biological and functions phenotypes [58]. This merit achieves from the same DNA sequence; cells might discriminate in actual dissimilar means. The renovation between euchromatin and heterochromatin is called epigenetic regulation (Perria et al. 2017).
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
