
Research Article | DOI: https://doi.org/10.31579/2640-1045/101
1Department of Radiology and Research, Veteran Administration University of California Irvine Medical Center, Long Beach, CA 90822.
2Department of Medicine, University of Massachusetts Medical School, Worcester, MA 01605.
3Department of Physiology and Anatomy, California State University of Long Beach, CA 90803.
4Department of Obstetrics and Gynecology, University of California Irvine, Irvine, CA 92697.
*Corresponding Author: Sing-yung Wu, Radiology and Research Service (151), VA-UCI Medical Center, Long Beach, CA 90822, USA.
Citation: Sing-y Wu, Charles H Emerson, E Tjioe, Dong B Chen . (2021). Maternal 3,3’-Diiodothyronine Sulfate Formation from Guinea Pig Placenta Perfused with 3,3’,5-Triodothyronine. Endocrinology and Disorders. 5(6): DOI:10.31579/2640-1045/101
Copyright: © 2021 Sing-yung Wu, This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 17 September 2021 | Accepted: 09 October 2021 | Published: 25 October 2021
Keywords: placental function; placental transfer; thyroid hormones; sulfation of thyroid hormone
Objective: Serum 3, 3’,5-triiodothyronine (T3) remains low in near-term fetus to prevent the growing fetus from undue exposure to its active catabolic effect in mammals. The present study was undertaken to gain insight in the role of placenta in T3 metabolism, fetal to maternal transfer of T3, and its metabolites by in situ placenta perfusion with outer-ring labeled [125I]-T3 in pregnant guinea pig, a species showing increased sulfated 3, 3’-diiodothyronine (T2S) levels in maternal serum in late pregnancy (term = 65 days), similarly to humans in pregnancy.
Materials and Methods: One-pass placenta perfusions performed on pregnant guinea pigs were studied between 58 - 65 days of gestation. In two separate experiments, the umbilical artery of the guinea pig placenta was perfused in situ at 37°C with outer-ring labeled [125I]-T3. Maternal sera and umbilical effluents were obtained for analysis at the end of a 60-minute perfusion, when the steady-state levels of radioactivity were reached in the placenta effluent after 30-minute.
Results: Sulfated [125I]-T2S was readily detected in the maternal serum as the major metabolite of T3 following the perfusion of placenta with [125I]-T3, suggesting that placental inner-ring deiodinase and sulfotransferase may play an important role in fetal T3 homeostasis and in the fetal to maternal transfer of sulfated iodothyronine metabolites.
Conclusions: The expression of type 3 deiodinase (D3) and thyroid hormone sulfotransferase activity in placenta may play an important role to protect developing organs against undue exposure to active thyroid hormone in late gestation in the fetus. The combined activities of D3 and sulfotransferase promoted a placental transfer of T2S into maternal circulation. The maternal circulation of T2S is fetal T3 in origin and its role as a fetal thyroid function biomarker deserves further evaluations and studies.
Optimal level of thyroid hormone (TH) is essential for normal neurological development in developing mammals, including humans, A critical amount of TH is especially important in the central nervous system (CNS) maturation. Deficiency or excess of TH in the CNS during fetal and neonatal periods can lead to morphological and functional abnormalities [2, 3]. The most severe form of TH deficiency in human fetus and neonate is the syndrome of cretinism. A milder form of fetal TH deficiency is observed in children of women with high serum TSH during pregnancy who performed less well in IQ and other neuropsychological tests [4, 5]. These findings underline the importance of TH in human brain development.
Thus, the study of fetal TH metabolism has been the subject of considerable interest. Iodothyronines detected in the fetus before the onset of fetal thyroid function must be maternal in origin [6]. The maternal-fetal transfer of TH and their metabolites is apparently a two-way street [1]. The high gradient between fetal and maternal serum concentrations of iodothyronine sulfates raises the possibility of significant fetal to maternal transfer of iodothyronine sulfoconjugates [1, 2]. Sack et al. [7] showed that umbilical cord cutting, thus removing the lamb from placental type 3 deiodinase (D3) and fetal-to-maternal transfer, triggers hypertriiodothyroninemia in the newborn lamb and that the postnatal T3 peak can be delayed until well after the TSH peak by delaying umbilical cord cutting. Santini et al. [8] found that the placenta plays an important role in maintaining the low serum T3 in fetuses late in gestation. These findings suggest the importance of the placenta in fetal T3 metabolism, and it is possible that fetal-to-maternal transfer of the sulfated iodothyronines via placenta is one mechanism responsible for optimization of serum T3 concentrations in the fetus. Increasing fetal-to-maternal transfer of iodothyronines may occur in late gestation [1].
Sulfoconjugation is the dominate pathway in thyroid hormone metabolism during intrauterine development, particularly in the last trimester in the precocial species, including humans, guinea pigs, and sheep. It regulates the supply of T3, via sulfation and deiodination, and facilitates fetal to maternal transfer of sulfated iodothyronines, e.g., 3,3’-diiodothyronine sulfate (T2S) [1, 9].
Sulfation, catalyzed by sulfotransferase enzymes (SULTs), is an important pathway of thyroid hormone metabolism, irreversibly converting T4 to inactive reverse T3 (rT3) rather than active T3. The human fetus and neonate have high levels of circulating sulfated iodothyronines (e.g., T4S, T3S, rT3S, and 3,3’-T2S); the placenta forms the link between the fetus and its mother and is involved in transfer of thyroid hormones early in pregnancy. We and others have examined the expression of the SULTs involved in iodothyronine metabolism during mammalian placental/uterus development [10, 11]. The placenta at late gestation may facilitate inactivation of T3 and fetal to maternal transfer of 3,3’-T2S. Thus, it is possible that T2S or its derivatives, transferred from the fetus and appearing in maternal serum or urine, can serve as a biomarker of fetal thyroid function; and in humans we have previously reported W-Compound to fill this role [1].
The aim of present study was undertaken to gain insight in the role of placenta in the fetal to maternal transfer of T2S by in situ placenta perfusion with outer-ring labeled [125I] -T3 in the pregnant guinea pig, a species showing increased T2S levels in maternal serum in late pregnancy (term = 65 days), similarly to humans [1]. We found that [125I]-3, 3’-T2S was readily detected in the maternal serum following the perfusion of placenta with [125I] -T3. This suggests that placental inner-ring deiodinase and sulfotransferase may play an important role in fetal T3 homeostasis and in the fetal to maternal transfer of sulfated iodothyronine metabolites. It is consistent with the hypothesis that transferred sulfate iodothyronine (3,3’-T2S) in maternal compartment may serve as a biomarker for fetal thyroid function.
T2S radioimmunoassay
3,3’-T2S and [125I]-T2S were prepared by the method of Eelkman-Rooda et. al., Mol and Visser [12, 13]. T2S was further purified and quantitatively recovered by reverse-phase high-pressure liquid chromatography (HPLC) with a preparative column, as described previously [14, 15].
The T2S radioimmunoassay (RIA) procedure was performed as reported previously [14, 15], with modifications as below. Serum samples (0.2 to 1.0 mL) were extracted with 2 vol 95% ethanol (final concentration, 63%) before assay. Since the extraction efficiency of T2S in serum exceeded 96% in preliminary experiments, final T2S concentrations were not corrected for recovery rate. The lower limit of detection of the assay was 3.3 fmol (2 pg), or 33.1 pmol/L in a 300 uL ethanol extract of serum [14]. Intra-assay variations were 1.9% to 9.1% and inter-assay variations were 6.0% to 19.5%, depending on the measured concentrations [15].
With respect to dose-responsiveness, we found close parallelism to the T2S standard curve in the range between 10-250 pg/tube [15]. In the measurement of serum T2S, 100 µL of ethanol extract was used, which usually contains 5 to 100 pg per tube, corresponding to a T2S of 15 to 300 ng/dL (or 0.248 to 4.96 nmol/L) in the serum sample [15].
In Situ Perfusion of Guinea Pig Placenta
Normal male and female guinea pigs (500 grams) were purchased from Elm Hill Breeding Labs (Chelmsford, MA), and mated in our laboratory (at Worcester). Plug dated pregnancies were studied between 58 -65 days of gestation. The animal use protocol was approved by Institutional Animal Care and Use Committee at the Medical School, University of Massachusetts, Worcester.
One-pass placenta perfusions were performed using a technique modified from Kihlstrom and Kihlstrom [16, 17]. Anesthesia was induced with an intramuscular injection (i.m.) of Ketamine/Rompun (70 mg Ketamine and 6 mg Rompun/100 grams body weight). A catheter of PE5O tubing was tied into the maternal carotid for sampling of maternal serum. Following maternal cannulation, a laparotomy was performed to expose a single fetus through a small uterine incision. Both uterus and placenta were left within the maternal abdominal cavity to minimize hemodynamic changes in the maternal circulation to the placenta. To facilitate cannulation, we induced the dilation of vessels by the topical application of 4% papaverine. The umbilical vein and one fetal artery were cannulated with PE6O polyethylene tubing. The remaining vessels leading to the fetus were tied off and the fetus was removed [17].
In two separate experiments, the umbilical artery of the guinea pig placenta was perfused in situ at 37°C with outer-ring labeled [125I]-T3. The constituents of the perfusion media were 0.14 nM [125I]-T3, 3% bovine serum albumin (BSA) in Krebs-Henseleit (KH) buffer (0.110 M NaCl, 2.4 mM CaCl2, 4.4 mM KC1, 1.1 mM KH2PO4, 1.1 mM MgSO4 7H2O, and 25.0 mM NaHCO3, pH 7.4). Placental pressure was monitored with a string-gauge transducer and catheter positions were adjusted to maintain a placental pressure between 20-40 mmHg. Maternal sera and umbilical effluents were obtained for analysis at the end of a 60-minute perfusion, when the steady-state levels of radioactivity were reached in the placenta effluent after 30-minute [17].
High-pressure Liquid Chromatography (HPLC): Identification of Iodothyronines
Iodothyronines were identified by HPLC as below [14, 15]. Radioactive metabolites were identified in maternal serum extracts of guinea pigs which were perfused in situ via the fetal placental artery with [125I]-T3. Six to ten milliliters of maternal samples, obtained at the end of the 60-minute perfusion were extracted with two volumes of 95% ethanol and subsequently lyophilized. The dried extracts were dissolved in one milliliter of H2O and purified with a LH-20 column as previously described [14]. The infusate and effluent samples were diluted appropriately with 0.025 N NaOH. After application to the HPLC μBondapak C18 column, the serum extract was eluted isocratically with a mixture (22:78 vol/vol) of acetonitrile and 0.02 M ammonium acetate, pH 4.0, at a flow rate of 2 ml/min. Radioactive peaks were identified by comparing their retention times to those of known synthetic iodothyronines eluted on HPLC under the same conditions. Radioactivity in serum extracts was expressed as percent of total activity of each sample (100%).
Sources of Materials
[125I]3,3’-T2 was prepared by radioiodination using the method described previously [18]. BSA, 1-ethyl-3-(3-dimethyl-aminopropyl) carbodiimide, iodomethane, and dimethyl-formamide were purchased from Sigma Chemical Co. (St. Louis, MO). Chlorosulfonic acid (99%) was purchased from Aldrich Chemical Co. (Milwaukee, WI). The Biochrom 1010 ODS preparative column was obtained from Regis (Morton Grove, IL). Papaverine hydrochloride was purchased from Eli Lilly Co. (Indianapolis, IN) PE50 and PE60 poly-ethylene tubings were obtained from Fisher Scientific Co. (Medford, MA). [125I]-T3 was purchased from New England Nuclear (PerkinElmer Inc., Boston, MA) with its specific activity ranged from 1,133 to 2,000 μCi/ug.
Statistical Analysis
In the current study, the serum sample sizes are small (10 and 4 in the pregnant and virgin group, respectively), we use “testing for difference between two means” [19]. The two-tailed hypotheses, can be proposed to ask whether, in the small population sampled, the mean (X1) of the first group is different from the mean (X2) of the second group. These hypotheses are commonly expressed in their equivalent forms: The t value for testing the hypotheses concerning the difference between two means is:
t = X1 – X2/Sx1-x2
(The difference between two means divided by the standard error of the difference between the means.)
Significance was defined as p < 0>
3, 3’-T2S concentrations were significantly elevated in third-trimester pregnant guinea pig sera [0.64 ± 0.19 nmol/L (n=10) vs. 0.15 ± 0.04 nmol/L (n=4) in virgin female guinea pigs, p < 0>
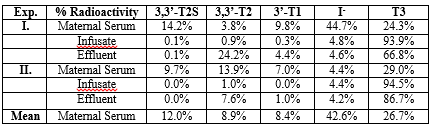
After perfusion of the near-term guinea pig placenta with [125I-T3] for 60 minutes using a reservoir system we demonstrated a distinct [125I]-T2S peak by HPLC in maternal serum extracts (Figure. 1). The T2S peak accounted for a mean of 12.0% of the total radioactivity in maternal serum extracts (Table 1). The remaining radioactivity consisted of 8.9% for 3,3’-T2, 8.4% for 3’-T1, 26.7% for T3 and 42.6% for free iodide suggesting a rapid inner-ring deiodination (Table 1). The sulfoconjugation appears to be a step to promote the placenta to maternal transfer of T3 metabolites.
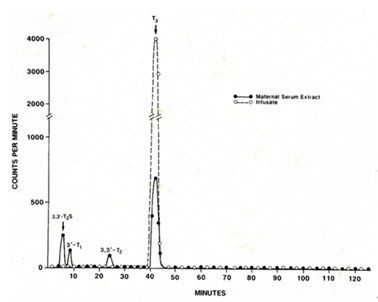
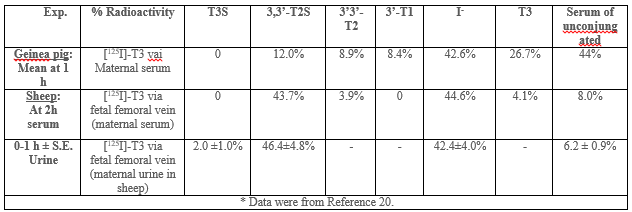
A comparison of [125I]-T3 infusion via the fetal placental artery in guinea pig (maternal serum) with via the fetal femoral vein in intact fetal lamb (maternal serum and urine in sheep) (20). Both guinea pig (58 – 65 d of gestation, term=65 d) and fetal lamb (139 – 143 d, term=150 d) are in near term. Table 2 shows much higher percent of T2S radioactivity is found the ovine serum, 43.7%, in the first two hours of infusion than the guinea pig, 12.0% in the first hour of maternal serum. The amount of free iodide is similar in the Sheep (42.6%) and guinea pigs (44.6%).
The expression of type 3 deiodinase (D3) in placenta/uterus, embryonic and fetal tissues may protect developing organs against undue exposure to active thyroid hormone (Figures. 2 and 3). Also in adult subjects, D3 appears to be an important site for clearance of plasma T3 and production of plasma rT3 and 3, 3’- T2 [1, 2, 14]. To remove placental D3 and fetal-to-maternal transfer, Sack et al. [7] showed that umbilical cord cutting triggers hypertriiodothyroninemia in the newborn lamb.
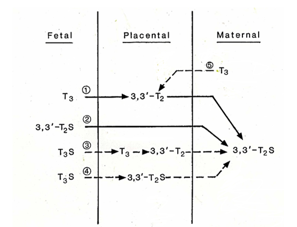

In rats, as in humans, sulfation of iodothyronines is catalyzed by multiple sulfotransferases (SULT) isozymes in different tissue [1, 9]. We have identified rSULT1A1 and 1B1 mRNA in rat uterus but not 1C1 [1]. In rats, we found significant activities in uterus with an apparent Km of 0.62 μM for T2, and activity varied during gestation [10]. In various substrates SULTs of iodothyronine, 3,3’-T2 is by far the best of substrate as compared to other thyroid hormone analogs: 3,3’-T2 > rT3 >> T3 >T4 [10]. From Figure. 1 and Table 1, we find 3, 3-T2S is the major metabolites in both intact fetal infusion in near-term fetal lamb [20] and in situ isolated fetal placental artery perfusion suggesting that T3 is rapidly degraded by D3 [21] to 3, 3’-T2 first and then sulfated to 3, 3’-T2S. SULT1A1 activity showed significant correlation with sulfation of 3,3'-T2, suggesting that this enzyme is primarily responsible for placental T2 sulfation. It is likely that SULT isozyme in guinea pig and sheep in placenta/uterus that resembles rSULT1A1 that has high affinity to 3,3’-T2.
We and others have reported high concentrations of sulfated iodothyronines, including thyroxine sulfate (T4S), T3 Sulfate (T3S), and reverse T3 sulfate (rT3S), in sera of human and sheep fetuses [22 – 25]. Levels of these metabolites in maternal sera are low. We found Increased Levels of T2S– immune-cross-reactive material was found in both fetal and maternal sera during human pregnancy [25, 26].
In sheep study, we have shown T2S derived from T3 of fetal origin is transferred to the maternal circulation and contributes significantly to the maternal urinary pool [1, 27 – 29]. We also have reported that T2S was the major metabolite in maternal compartment while labeled T4 or T3 was infused intravenously to fetuses in utero at late gestation [20, 28]. These findings suggest the importance of the placenta in fetal T3 metabolism, and it is possible that placental fetal-to-maternal transfer of the sulfated iodothyronines is one mechanism responsible for optimization of serum T3 concentrations in the fetus. Thus, fetal-to-maternal transfer of iodothyronines may increase in late gestation.
In the present study, we have observed that when the umbilical artery of the guinea pig placenta was perfused in situ with outer-ring labeled T3 and 3, 3’-T2S was found to be the major product in maternal compartment. Again, this finding is consistent with the maternal circulation of 3,3’-T2S, in mammalian species, or its immuno-crossreactive material (W-Compound) is fetal T3 in origin and could be used a fetal thyroid function biomarker [1, 26, 30, 31].
The limitations of the present study include that it is a small study with only two perfusions, no statistical analysis could be performed. In addition, no specific manipulations of D3 or SULTs with inhibitors are performed. SULT activity may be inhibited by xenobiotics [32]; D3 can be inhibited by propylthiouracil (PTU) [33]. Further studies with specific inhibitors may differentiate the role of D3 and SULTs in placental metabolism and transfer of iodothyronines. Finally, future study with in vitro perfused human placental lobule, that is collected and prepared immediately after delivery from normal uncomplicated pregnancy, may be more relevant to humans [34].
Recently, brominated flame retardants (BFRs) have been shown to disrupt thyroid hormone (TH) homeostasis through multiple mechanisms, including inhibition of enzymes that regulate intracellular levels of THs, such as sulfotransferases (SULTs). As shown in the present study, the placenta plays a critical role in expressing D3 and SULTs to prevent the developing fetuses from exposure to high level of active thyroid hormone T3, which is needed immediately after birth. This is concerning given that disruption of TH regulation within the placenta could potentially harm the developing fetus [35]. The possibility of the maternal serum or urine levels of W-Compound also be used as a marker for BFR toxicity may be explored.
The expression of type 3 deiodinase (D3) and thyroid hormone sulfotransferase activity in placenta may play an important role to protect developing organs against undue exposure to active thyroid hormone in late gestation in the fetus. The combined activities of D3 and sulfotransferase promoted a placental transfer of T2S into maternal circulation. This finding is consistent with our previous studies in human and sheep that the maternal circulation of 3,3’-T2S, in mammalian species, or its immuno-cross-reactive material (W-Compound) is fetal T3 in origin and its role as a fetal thyroid function marker deserves further evaluations and studies.
This work has been supported by the Department of Veterans Affairs, NIH RO1 HL70562 and NIH R21 HD097498.
None.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
