
Review Article | DOI: https://doi.org/10.31579/2692-9406/106
1 International Higher School of Medicine, Intergelpo str, 1F, Bishkek, Kyrgyzstan.
2 National Center of Cardiology and Internal Medicine, Togolok Moldo str,3, Bishkek, Kyrgyzstan.
*Corresponding Author: Abyt Ibraimov. International Higher School of Medicine, Intergelpo str, 1F, Bishkek, Kyrgyzstan.
Citation: Ibraimov A, Akhunbayev S and Uzakov O. (2022). Human Chromosomal Q-heterochromatin Regions as a System. Biomedical Research and Clinical Reviews. 6(4); DOI: 10.31579/2692-9406/106
Copyright: © 2022 Abyt Ibraimov, This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 01 February 2022 | Accepted: 26 February 2022 | Published: 04 March 2022
Keywords: constitutive heterochromatin; chromosomal Q-heterochromatin regions; epigenetics; cell thermoregulation; genetic system
The eukaryotic genome consists of the two forms of chromatin: euchromatin and heterochromatin. The DNA of euchromatin contains the gene portion of the genome, while heterochromatin is represented predominantly from non-coding repetitive nucleotide sequences that do not encode proteins or enzymes. In higher eukaryotes, some part of the non-coding, highly repetitive nucleotide sequences were transformed into complex forms of DNA organization as chromosomal constitutive heterochromatin regions. There are two types of constitutive heterochromatin: C- and Q-heterochromatin. C-heterochromatin is found in the chromosomes of all eukaryotic cells, while Q-heterochromatin is found in the karyotype of only three higher primates (Homo sapiens, Gorilla gorilla and Pan troglodytes).
Since the discovery of the position effect variegation phenomenon C-heterochromatin has been attributed to gene silencing effects. Dosage compensation of genes is another epigenetic gene silencing mechanism that makes it possible to equalize the level of expression of sex-linked genes in males and females. In mammals, this is done by inactivating one X chromosome in the cells of females using facultative heterochromatin, which is a heterochromatinized euchromatin. However, no epigenetic gene silencing was found in chromosomal Q-heterochromatin regions (Q-HRs). The question is discussed why human chromosomal Q-HRs does not exhibit gene silencing or other epigenetic effects and what their biological role might be.
The two forms of chromatin were distinguished by cytological staining of cells: heterochromatin and euchromatin. Heterochromatin stained darker than euchromatin due to a denser packing and is generally localized to the periphery of the nucleus. The DNA of euchromatin contains the gene portion of the genome, while heterochromatin is represented predominantly from non-coding repetitive nucleotide sequences that do not encode proteins or enzymes. In higher eukaryotes, some part of the non-coding, highly repetitive nucleotide sequences were transformed into complex forms of DNA organization as chromosomal constitutive heterochromatin regions (HRs). There are two types of constitutive heterochromatin: C- and Q-heterochromatin. C-heterochromatin is found in the chromosomes of all eukaryotic cells, while Q-heterochromatin is found in the karyotype of only three higher primates (Homo sapiens, Gorilla gorilla and Pan troglodytes).
Position effect variegation (PEV) causes a gene to be stochastically silenced in regions where it should normally be expressed. This occurs when the gene becomes juxtaposed to C-heterochromatin in a genome translocation or insertion event. An example is variegated eye color in Drosophila caused by a translocation of the white gene into the vicinity of heterochromatin.
Dosage compensation of genes is an epigenetic mechanism that makes it possible to equalize the level of expression of sex-linked genes in males and females of those species in which sex determination is carried out using sex chromosomes. In mammals, this is done by inactivating one X chromosome in the cells of females, so that in each somatic cell of an individual of either sex, there is only one active X chromosome per diploid set of chromosomes.
Chromosomal Q-HRs variability in human population.
It is known that chromosomal C-heterochromatin regions (C-HRs) are present on all 46 chromosomes in the human karyotype and their complete absence even in one chromosome is extremely rare. It has been established that the C-HRs makes up 15-29% of the human genome. Unlike C-HRs, chromosomal Q-heterochromatin regions (Q-HRs) may be completely absent in the human genome without any noticeable pathological or other phenotypic manifestations for the carrier. According to some authors, individuals with a complete absence of Q-HRs make up a significant part of human populations [1-8]. There is a huge literature, including a number of reviews devoted to the study of morphology, molecular biology and methods of identification of chromosomal C- and Q-HRs in the human karyotype [6,9-16]. The main morphological manifestation of morphological variability of Q-HRs in the human karyotype is the presence, degree of fluorescence, size and location of Q-heterochromatin segments in 12 potentially Q-polymorphic loci of seven autosomes (3p11q11, 4p11q11, 13p11, 13p13, 14p11, 14p13, 15p11, 15p13, 21p11, 21p13, 22p11 and 22p13) and in the q12 segment of the Y chromosome. It should be specially emphasized that not a single individual was found in the human population in whose karyotype Q-HRs was present in all 25 potentially Q-polymorphic loci. In the human population, the number of Q-variants usually ranges from 0 to 10 [3-6,17-20].
Currently, the following quantitative characteristics of the variability of Q-heterochromatin polymorphism are used to estimate the amount of chromosomal Q-HRs in the genome of a human population: 1) the frequency of Q-HRs in 12 potentially polymorphic loci of seven autosomes, which are usually expressed as a percentage from the number of analyzed chromosomes; 2) the distribution of the numbers of Q-HRs in the population, i.e. the distribution of individuals with a different amount of Q-HRs regardless of their localization in the karyotype, which also reflects the magnitude of the variability of the amount of Q-HRs in the sample; 3) the mean number of chromosomal Q-HRs per individual in the population (m), which is determined by dividing the total amount of Q-HRs found in this sample by the number of studied chromosomes; and 4) the size of the Y chromosome, which is conventionally divided into three groups: (a) large (Y = F), (b) medium (F > Y > G) and (c) small (Y = G) [15,21].
Suggestions on the possible biological role of the constitutive heterochromatin.
There are many hypotheses regarding the possible biological role of constitutive C-heterochromatin [9-15]. However, apart from the PEV phenomenon, almost all of the hypotheses put forward have not yet been experimentally confirmed. Once again, we specifically note that PEV was found in animals that lack chromosomal Q-HRs in the genome. In species with chromosomal Q-HRs - Homo sapiens, Pan troglodytes and Gorilla gorilla - phenomena directly or indirectly resembling PEV phenomenon have not been described.
There is still no consensus on the nature of the variability of chromosomal Q-HRs in the human genome, although all hypotheses adhere to the "selectionist" approach. Only one of these hypotheses implies that for Q-HRs, “the dose, and not the localization of Q-HRs on a particular chromosome, is of paramount importance” [8,21]. Here the term “dose" refers to the total amount of Q-heterochromatin material in an individual's genome. In other words, this hypothesis is based on the assumption that chromosomal Q-HRs do not possess locus specificity.
All other hypotheses hold the point of view that "localization is important for an individual, not the amount of Q-HRs”, that is, they are supporters of a locus-specific approach. Proponents of this approach believe that, for example, deviations from the theoretically expected Q-HRs frequencies at the population level reflect some structural and functional features of these loci and are due to either selection or non-random segregation of chromosomes carrying Q-HRs [22-24]. They suggest that Q-HRs localized on different chromosomes in the human karyotype differ in structure and biological effects. This, they believe, is evidenced by deviations in the observed frequencies of homo (+/+ or -/-) and heterozygotes (-/+) of chromosomal Q-HRs from those theoretically expected when they are checked for compliance with the Hardy-Weinberg law (for more details see [15]).
On this occasion, Ginzburg et al. [25] specially studied the statistical side of this controversial issue. Leaving aside the question of the validity of the interpretation of the violation of the Hardy-Weinberg rule, the authors focused on the statistical procedure by which this violation can be detected. The authors showed that if the χ2 criterion is used for small samples (often the volumes did not exceed 25-200 observations), and the Hardy-Weinberg distribution hypothesis is respected, the researcher, nevertheless, will reject it more often, and, in some cases, the error of the first kind reaches a value that makes the study itself incorrect.
Thus, it can be assumed that the use of non-correspondence of the observed frequencies of homo- and heterozygotes to those theoretically expected in some samples as an argument in favor of the hypothesis of locus specificity of chromosomal Q-HRs is not justified. This method, being formal in nature, did not advance in understanding the biological role of the wide variability of Q-HRs in human populations. This idea was most clearly expressed by Jacobs [26], who noted that it is premature to assess the correspondence of the observed and expected frequencies of homo- and heterozygotes to the Hardy-Weinberg law, since existing methods for quantifying chromosomal Q-variants are far from perfect due to the continuous nature of the distribution of Q-HRs in size in human populations. Mayr [27] specifically studied this question and came to the conclusion: “They expressed this as a mathematical formula, which is a reapplication of a mathematical law, the binomial expansion. Being a strictly mathematical solution, it is not a biological law”.
Our attitude to this issue is as follows: until evidence is obtained that Q-HRs localized in 12 potentially polymorphic loci of seven pairs of autosomes and on the human Y chromosome are fundamentally different structures in molecular and/or genetic terms, the use of the Hardy-Weinberg rule remains mainly an exercise in solving some problems of applied mathematics without significant biological content. On the whole, it can be assumed that observations of supposedly deviations in the segregation of Q variants in families or a discrepancy between the observed and theoretically expected frequencies of homo- and heterozygotes in some samples do not yet require a special biological interpretation [15,28].
Our assumption about the absence of locus specificity of human chromosomal Q-HRs is based on numerous observations that with an increase or decrease in the mean number of Q-variants per individual in the population, a proportional increase or decrease in the frequency of Q-variants occurs for all Q-polymorphic loci [15,18,29]. In particular, if we arrange the studied samples according to the value of the mean number of Q-HRs per individual and perform the same operation with the frequencies of Q-variants on seven Q-polymorphic autosomes, then the distribution of populations by the frequencies of Q-variants in these autosomes will correspond to their distribution by the value of the mean numbers of Q-HRs calculated per person in the population (Table 1). A similar result was obtained by us after analyzing the data of other researchers [30-32], which revealed the existence of inter-population differences in the values of the mean number of Q-HRs [15,29,33].
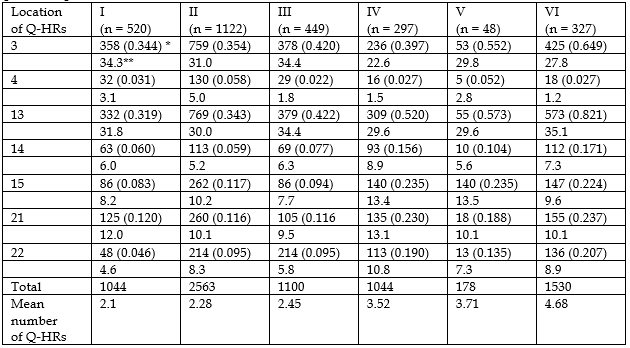
The totality of the data presented below shows the validity of the assumption that there is no locus specificity of chromosomal Q-HRs: a) despite the fact that there are 25 loci in the human karyotype in which may be chromosomal Q-HRs, in fact, the maximum number of Q-variants does not exceed ten [1,2,6]; b) in human populations, the number of Q-HRs usually ranges from 0 to 10 without visible pathological or other phenotypic manifestations [6]; c) the distribution of Q-variants in the samples has always been close to the normal distribution [18,21,34 ]; d) at the population level, the distribution of Q-HRs on seven Q-polymorphic autosomes is uneven: the largest number of Q-variants was found on chromosomes 3 and 13 (more than 50%), the remaining Q-HRs were more or less evenly distributed on the remaining five autosomes [29,34] (see Table 1); e) human populations do not differ from each other in the relative content (“portion”) of Q-HRs on seven autosomes: the portion of Q-variants on autosomes 3, 4, 13, 14, 15, 21, and 22 were on average, 25.5 %, 3.5 %, 30.7 %, 8.6 %, 12 %, 10.6 %, and 9.1%, respectively [29,33] (see Table 1); f) the quantitative content of Q-HRs in the genome of a human population is best determined by the mean number of Q-variants per individual (m) in the sample [1-6,8,21,34]; g) significant differences were found between the samples in the quantitative content of chromosomal Q-HRs, and it turned out that these differences are related to the climatic and geographical features of the place of permanent residence of the studied populations, and not to their racial and ethnic composition [3-6,18-21,34]; h) changes in the mean number of Q-HRs in the genome of populations tend to decrease from low to high geographical latitudes and from low to high altitudes [3-6,15]; i) both a decrease and an increase in the value of m is accompanied by a narrowing or expansion of the range of variability of Q-variants [29,33]; j) in different age groups, the values of m differ — the largest number of Q-variants are present in the genome of newborns, and the smallest in individuals from older age groups [30,36]; k) individuals who adapted to hypoxic hypoxia at high altitudes (mountaineers) and the extreme climate of the Far North of Siberia had an extremely low amount of chromosomal Q-HRs in the karyotype [3-6,18-21,34]; l) Q-HR on the Y chromosome, being the largest block of Q-heterochromatin in the human karyotype "restricts" the amount of Q-HRs on autosomes, namely, in samples of men with large Q-HR sizes on the Y chromosome, the m value is significantly lower and vice versa [37]; m) there is some evolutionarily fixed mechanism that compensates for the low content of Q-heterochromatin in the female genome due to the absence of a Y chromosome in their karyotype by increasing the amount of Q-HRs on their autosomes [38]; and, finally, n) individuals in the human population differ from each other in the level of thermal conductivity of their bodies, and these differences are related to the amount of chromosomal Q-HRs in their genome [39].
Thus, more than half a century of experience in studying chromosomal Q-HRs shows that this type of constitutive heterochromatin has not demonstrated effects even remotely resembling PEV, gene silencing or other known epigenetic processes. This is directly evidenced, at least, by the fact that even the complete absence or maximum amount of Q-heterochromatin in the human karyotype has no visible phenotypic manifestation, so that they could be used to judge the effect of Q-HRs on the functioning of genes. On the other hand, chromosomal Q-HRs at the level of human populations behave so regularly that their behavior can be predicted.
Human chromosomal Q-HRs as a system.
Now let's consider an approach in which human chromosomal Q-HRs are considered as a single structural and functional system, where their total amount (dosage) in the genome is of primary importance, and not localization on one or another chromosome in the karyotype. If it concerns a concrete person, then an important criterion for assessing the variability of Q-variants should be their total number (from 0 to 10) in the karyotype; when it comes to a population, then the mean number of Q-HRs per individual in the population is crucial [15,18].
Usually, by "system” it is customary to mean a certain totality, which in some way can be distinguished from real or imaginary elements of the surrounding world [40]. In biology, consider a particular set as a system if: 1) each of its elements is indivisible (not in the physical sense, but conditionally); 2) it interacts with the surrounding world as a whole; and 3) quantitative relations are established between the elements of the selected set, and in its evolution, correspondences are preserved between the elements, that is, there is some ordering.
The indivisibility of chromosomal Q-HRs located in 12 polymorphic loci of seven pairs of autosomes and Y chromosomes is evidenced by the specificity of cytochemical staining methods (Q-staining) used to detect them [41]. The second requirement meets such facts: an increase or decrease in the frequency of human chromosomal Q-HRs occurs simultaneously across all Q-polymorphic loci in proportion to their initial frequencies in the population, regardless of gender, age, racial and ethnic characteristics or climatic and geographical features of their places of permanent residence. If the elements of the proposed system should interact with the surrounding world as a whole, then a proportional decrease or increase in the number of Q-variants for all Q-polymorphic autosomes in the genome of a population, depending on the characteristics of the ecological environment of their habitat, may indicate in favor of considering chromosomal Q-HRs as a single structural and functional system (see also Table 1) [15,18,33].
Regarding the third requirement, it should be noted that: a) the proportion of Q-HRs variants in 12 Q-polymorphic loci of seven pairs of autosomes in human populations, expressed as a percentage of the number of Q-variants are always comparable, regardless of the average values of the number of Q-HRs in the samples [15,29,33]; b) even in those populations where the value of the average number of Q-HRs is very low, Q-heterochromatin does not disappear on autosomes that differ with low frequency, and moreover, the contribution of Q-highly polymorphic and Q-low polymorphic chromosomes in the total pool of Q-HRs in the samples remains comparable [3-6,18-21,34]; c) the distribution of Q-variants in the population is always close to the normal distribution [15,29]. All this testifies in favor of the point of view that chromosomal Q-HRs behaves at the level of human populations as a single self-sustaining structural and functional system [15,21,29].
When asked why chromosomal Q-HRs behave as a single genetic system and, contrary to theoretical expectations, do not show any signs of epigenetic effects, we would answer as follows. The consistency in the behavior of human chromosomal Q-HRs at the population level is due to their special biological role. We believe that chromosomal Q-HRs, together with C-HRs, are involved in thermoregulation at the cell level, the essence of which is that they, as part of a dense layer of condensed chromatin (CC), contribute to the dissipation of excess heat from the interphase nucleus.
DNAs of human chromosomal constitutive HRs are not involved in the production of proteins, enzymes, or RNAs, and their main biological effect is the dense packing (compactization) of the CC layer around the interphase nucleus. This CC layer, being the densest structure in the interphase cell, must have a high thermal conductivity in order to facilitate more efficient removal of excess heat from the nucleus.
The phenotypic manifestation of the biological effect of constitutive chromosomal HRs is the level of thermal conductivity of the human body, which has received experimental confirmation. In particular, our studies indicate that the phenotypic effect (the level of heat conductivity of the human body) of Q-HRs depends on their amount in the individual's genome, and not on the peculiarity of their localization on one or another chromosome in its karyotype. Apparently, the total number of human chromosomal Q-HRs in the genome of populations is under the control of natural selection, the pressure of which is determined by the climatic and geographical features of the environment where the population permanently lives (for details see [15,39,41]).
The eukaryotic genome consists of the two forms of chromatin: euchromatin and heterochromatin. The DNA of euchromatin contains the gene portion of the genome, while heterochromatin is represented predominantly from non-coding repetitive nucleotide sequences. There are two types of constitutive heterochromatin: C- and Q-heterochromatin. C-heterochromatin is found in the chromosomes of all eukaryotic cells, while Q-heterochromatin is found in the karyotype of only three higher primates (Homo sapiens, Gorilla gorilla and Pan troglodytes).
Since the discovery of the position effect variegation phenomenon C-heterochromatin has been attributed to gene silencing effects. Dosage compensation of genes is another epigenetic gene silencing mechanism that makes it possible to equalize the level of expression of sex-linked genes in males and females. However, no epigenetic gene silencing was found in chromosomal Q-heterochromatin regions (Q-HRs).
Our studies indicate that chromosomal Q-HRs behave as a single genetic system and do not show any signs of epigenetic processes. We believe that the systemic nature in the behavior of human chromosomal Q-HRs at the population level is associated with their special biological role. Apparently, chromosomal Q-HRs together with C-HRs are involved in thermoregulation at the cell level, the essence of which is that they, as part of a dense layer of condensed chromatin, contribute to the dissipation of excess heat from the interphase nucleus with all the ensuing consequences.
I apologize to that author whose work is not cited or is cited only through reviews. The reason for this is only the space limitations of the publication.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
