
Mini Review Article | DOI: https://doi.org/10.31579/2690-1919/371
1 Laboratory of Immune Regulation, The World Premier International Research Center Initiative (WPI) Immunology Frontier Research Center, Osaka University, 565-0871 Osaka, Japan.
2 Biochemistry Department, Biotechnology Research Institute, National Research Center, P.O. 12622, Giza, Egypt.
*Corresponding Author: Hozaifa Metwally, Tadamitsu Kishimoto, The World Premier International Research Center Initiative (WPI) Immunology Frontier Research Center, Osaka University, Japan.
Citation: Maha M. Elbrashy, Hozaifa Metwally and Tadamitsu Kishimoto, (2024), Gut Microbiome Metabolites and the Intestinal Homeostasis,
J Clinical Research and Reports, 15(4); DOI:10.31579/2690-1919/371
Copyright: © 2024, Hozaifa Metwally and Tadamitsu Kishimoto. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 22 April 2024 | Accepted: 09 May 2024 | Published: 16 May 2024
Keywords: gut microbiota; intestinal homeostasis; metabolites
Gut microbiota, the collection of microorganisms harboring the gut of all mammals, are essential for the host’s physiology, metabolism, intestinal homeostasis and immune system development and function. The advancement of technology has allowed a better understanding of the gut microbial communities and their metabolites, how they influence the host’s physiology and homeostasis, and how they also influence the susceptibility of the host to many diseases and disorders.In this mini - review, we will discuss how the intestinal microbiota metabolites regulate gut barrier function, epithelial proliferation, balanced gut microbiota composition, and intestinal immune activation.
The mammalian intestine harbors a diverse and active community of trillions of microorganisms (1). The dynamic host-microbiota interaction is important for the host’s physiology and homeostasis. Maintaining mutualistic symbiotic host-microbiome relationship allows the host to adapt diverse saccharolytic enzymes that complement the limited saccharolytic diversity encoded in the mammalian genome (2). Coordinated crosstalk among microbiota, and the host’s intestinal epithelial and immune cells ensure intestinal homeostasis and avoid abberant immune activation (3,4). Perturbation of the symbiotic host- microbiome interaction leads to dysbiosis -an imbalance of the composition or function of microbiome-, which is associated with inflammatory bowel diseases (IBD) such as Crohn’s disease (CD) and ulcerative colitis (UC) (5,6). Recent genome-wide association studies and animal models suggest that dysfunction of tight junctions (TJ) in the intestine is associated with the pathogenesis of IBD (5,7). Moreover, dysbiosis is associated with alterations in allergies (8), metabolic disorders (9), and immune-related diseases (10).
Dysbiosis is caused by environmental factors such as a changes of the diet and antibiotics that may destroy the epithelial barrier function and initiate a proinflammatory response (11). The gut microbiome is crucial for the metabolism of diverse dietary compounds to produce active metabolites that provide information to the host about the microbiome composition, the existence of pathogens, or other environmental stress (12). With the advancment of metabolomics analysis technologies, it has become available to observe changes of different types of metabolites in IBD patients in comparison with healthy subjects, such as bile acids, medium-and short- chain fatty acids, and polyamines (13). Numerous studies have reported that the impact of the gut microbe metabolites on gut barrier function (14). Besides, there are clinical trials using microbiome bioactive metabolites to restore the healthy intestinal barrier function in IBD patients (15–17).
In this mini - review, we will discuss how the intestinal microbiota metabolites regulate gut barrier function, epithelial proliferation, balanced gut microbiota composition, and intestinal immune activation.
Short-chain fatty acids
The human diet is rich in dietary fiber and indigestible carbohydrates that cannot be digested in the proximal digestive system. Instead, they are fermented by gut commensals, including Bifidobacterium, Bacteroides, Enterobacter, Faecalibacterium, and Roseburia species, which produce short-chain fatty acids (SCFAs) as a byproduct (18). SCFAs mainly comprise acetate, propionate and butyrate (19,20). SCFAs impact gut microbiota composition and the host’s metabolism and immune responses (21). Abberant levels of SCFAs are linked to disease development such as diabetes, obesity, and colorectal cancer (22).
SCFAs boost TJ protein expression, paracellular transport, zonula occludens (ZO)-1 and occludin (OCLN) maintenance, inhibit NLR family pyrin domain containing 3 (NLRP3) inflammasome activation, and enhance autophagy (23). In addition, high concentrations of SCFA enhances anion release, which maintains gut microbiome symbiosis (24,25). Diets rich in SCFAs promote colonic mucin-2 (MUC2) and OCLN expression, enhancing gut barrier function, and butyrate may reduce inflammation-induced barrier disruption (26,27). Furthermore, SCFAs regulate intestinal barrier by stimulating the secretion of antimicrobial peptides and secretory immunoglobulin A (sIgA), which prevent the adherence and invasion of harmful bacteria (28). Butyrate serves as the primary energy source for colonocytes (29), with human colonocytes from different regions utilizing over 70% of oxygen through butyrate oxidation (30). Patients with IBD exhibit dysbiosis and reduced levels of SCFAs/butyrate-producing bacteria (21).
Bile acids
Bile acids (BAs) are synthesized in its primary form by pericentral hepatocytes, followed by their transfer to the gut via enterohepatic circulation (31). The conversion of primary BAs into their secondary form is executed by the gut microbiota such as Bifidobacterium, Bacteroides, Clostridium, Lactobacillus, Enterobacter (32,33). In humans, primary bile acids include cholic acid (CA) and chenodeoxycholic acid (CDCA) converted into secondary bile acids such as deoxycholic acid (DCA), ursocholic acid, ursodeoxycholic acid (UDCA), and lithocholic acid (LCA). In mice the primary bile acids are cholic acid, CDCA, 6- hydroxylated bile acids, and muricholic acid, which further converted into the secondary bile acids DCA, ursocholic acid, UDCA, murideoxycholic acid, and hyodeoxycholic acid (34).
Secondary BAs exert multiple biological functions on the intestine via binding to the nuclear membrane farnesoid X receptor (FXR) and the G protein–coupled receptor; Takeda G protein-coupled receptor 5 (TGR5) in the intestines (35). In human epithelial cell line, Caco-2, modification of the composition of BAs alters the permeability of the intestinal mucosa and impact the function of the barrier by regulating the expression of tight junction proteins (36). For example, mice undertaken high fat in their diet have an increse in DCA, which is a major risk factor enhancing colorectal cancer (37). UDCA supplementation has been shown to attenuate inflammation and reduce intestinal permeability caused by multidrug- resistant extended-spectrum β-lactamase (ESBL)-producing E. coli in colibacillus diarrhea of newborn dairy calves (38).
Furthermore, BAs modulate immune responses in the intestine (39). Two derivatives of LCA, 3-oxoLCA and isoalloLCA, suppress T helper 17 (Th17) cell differentiation and increased the differentiation of Treg cells through the production of mitochondrial reactive oxygen species (40). In addition, BAs impact the function of macrophages and affect the homeostasis of the intestinal immune barrier. High-fat diet impacts bile acids (e.g., increased CA and DCA), gut microbiota composition, pro- inflammatory macrophage polarization, which subsequently leads to intestinal inflamamtion (41,42).
Polyamines
Polyamines regulate multiple cellular functions including cell development, amino acid and protein synthesis, oxidative DNA damage, proliferation and differentiation (43). The natural polyamines, including spermidine, spermine, and their precursor putrescine are important for the host’s physiology, mainly for intestinal maturation and the differentiation and development of immune system (33,44,45). Gut microbes such as E. coli, Bacteroides spp. and Fusobacterium spp. can synthesize polyamines (46). Multiple studies have shown that polyamines are essential for maintaing the intestinal barrier through enhancing the expression of TJs and adherens junctions (AJs) (47–50). Additionally, polyamines suppress gut mucosal inflammation by inhibiting inflammatory cytokine synthesis in macrophages (51,52).
Tryptophan and indole derivatives
Gut micribiota such as Clostridium spp., Bacteroides spp. and Escherichia coli metabolize dietary tryptophan into indole and indole derivatives (53). Indole derivatives include indole acrylic acid, indole-3- aldehyde, indole-3-propionic acid, indole-3-acetic acid, and indole-3-acetaldehyde. Tryptophan metabolites function as bacterial quorum sensors to communicate with bacteria and their host. Besides, tryptophan metabolites mediate the host’s intracellular signaling via binding to aryl hydrocarbon receptor (AhR), pregnane X receptor and retinoid-related orphan receptor gamma-t (54,55).
Tryptophan metabolites’ signaling through AHR impact on the intestinal homeostasis by modulating immune responses through acting on Th17 cells, macrophages, dendritic cells, and neutrophils and also decrease the pathogen colonization (56,57). AhR signaling, primarily on immune cells, promotes the expression of interleukin-22 (IL-22), which maintains gut barrier integrity through promoting mucosal wound-healing and the expression of antimicrobial peptides (AMPs) by intestinal epithelial cells (57).
Choline metabolites
Trimethylamine (TMA) is a byproduct generated by the gut microbiota, such as Bifidobacterium, Firmicutes, Proteobacteria, Actinobacteria, Faecalibacterium prausnitzii from dietary amines such as choline, betaine, and carnitine that originate from animal-based foods by the action of hepatic flavin-containing monooxygenases (FMOs) such as FMO1 and FMO3 (33,58,59). TMA is absorbed into the portal circulation and is oxidized by the liver into trimethylamine-N-oxide (TMAO) (59,60). TMAO has been shown to display pro-inflammatory, pro-atherogenic, and pro-thrombotic properties (61–65). For example, it was reported that increased levels of TMAO promote oxidative stress and inflammation of endothelial cells, activate the inflammasome and inflammatory signaling in vascular smooth muscle cells, promote the transformation of macrophages into foam cells, promotes platelet hyperreactivity, and alters cholesterol transport and bile acid synthesis (66). TMAO may promote the pathogenesis of IBD by impacting autophagy and activating NLRP3 inflammasome (67).
Polyphenolic derivatives
Dietary polyphenols are metabolized by the gut microbiota such as Bifidobacterium, Lactobacillus, C. difficile, F. prausnitzii, to bioactive metabolites including urolithins (Uro A,B,C,D, and iso-urolithin A) (33,68,69). UroA enhances gut barrier integrity through upregulation of TJ proteins (70). Treatment with UroA alliviates colitis in mice through enhancing gut barrier function and supressing the production of pro- inflammatory cytokines (70).
Conclusion and future perspectives
The current mini review postulates a summary about the role of gut microbiome metabolites in the maintenance of the intestinal homeostasis through regulating the intestinal epithelial integrity, gut microbiota composition, epithelial proliferation, and intestinal immune homeostasis (Figure. 1). Metagenomic research on gut microbiota from IBD patients have revealed that gut microbiota actively interacts with IECs for the regulation of barrier integrity. Moreover, IBD patients is characterized by gut dysbiosis, which is associated with impaired intestinal barrier and aberrant immune activation. Better understanding of host-gut microbiome interaction will eventually become a valuable tool for harnessing the power of microbial metabolites to restore gut barrier integrity, and can reveal advanced therapeutics not only for intestinal inflammation, but also for other immune- and metabolic-related disorders.
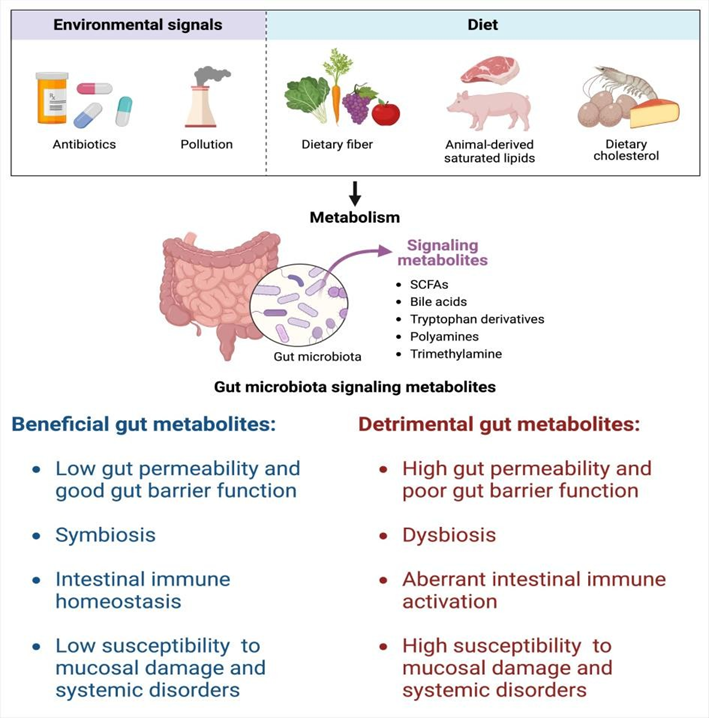
Figure 1. The role of gut microbiota metabolites in the regulation of intestinal homeostasis. Please explain the figure in the legend.
Environmental factors such as antibiotics and changes of the diet affect the balance of gut microbiota metabolites. Effects of beneficial (left) and detrimental (right) gut microbiota metabolites on intestinal homeostasis and inflammation.
M.M.E wrote the manuscript draft. H.M. and T.K edited the manuscriptand supervised the work.
Funding
This work was supported by Kishimoto Foundation. M.M. E. is supported by a full scholarship (ID: 69) from the Ministry of Higher Education of the Arab Republic of Egypt.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
