
Research Article | DOI: https://doi.org/10.31579/2692-9562/120
Family Physician & Public Health Consultant Bengaluru, India,
*Corresponding Author: Massoud AttarianShandiz, Independent Researcher; No. 18, Haft e Tir 40; Mashhad, Iran.
Citation: Massoud AttarianShandiz, (2024), Exploring the Impact of Berry Compounds on Virulence Gene Expression in Staphylococcus aureus to Shift Antimicrobial Strategies: A Microbiological Study, Journal of Clinical Otorhinolaryngology, 6(3); DOI:10.31579/2692-9562/120
Copyright: © 2024, Massoud AttarianShandiz. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
Received: 04 April 2024 | Accepted: 16 April 2024 | Published: 22 April 2024
Keywords: chronic sore throat; etiology; chronic tonsillopharyngitis; gastroesophageal reflux; laryngopharyngeal reflux
Staphylococcus aureus, a prevalent human pathogen, poses diverse health risks, ranging from mild skin conditions to severe illnesses, including potential foodborne infections and toxin production. This study investigates the influence of various berry extracts on the expression of key virulence genes (hla, spa, RNAIII) in S. aureus, aiming to evaluate their role as natural modulators of bacterial pathogenicity for combatting S. aureus infections. The examination focuses on natural compounds capable of reducing virulence.
Methods/aims:
Berry extracts were systematically tested against S. aureus to assess their impact on the expression of vital virulence genes under diverse growth conditions. Compounds influencing gene expression were identified through color changes observed on the plates. Interpretation of the plates relied on the emergence of a blue color, where high gene expression manifested as blue, while the absence of blue (appearing as white) indicated down-regulation.
Results:
Aronia, blackcurrant, blueberry, cranberry, and raspberry extracts consistently demonstrated the downregulation of hla, spa, and RNAIII genes. Notably, the spa gene exhibited the highest degree of inhibition. Additionally, anthocyanins extracted from berries displayed a significant reduction in spa gene expression.
Conclusions:
Berry extracts exhibit promising potential in mitigating S. aureus pathogenicity by repressing key virulence genes, with a pronounced effect on the spa gene. These findings offer valuable insights into the development of natural anti-virulence strategies for both therapeutic applications and advancements in food safety and preservation.
Staphylococcus aureus is a versatile pathogen capable of adapting to diverse environments within the human host. It is a Gram-positive bacterium known to cause a wide range of infections in humans, ranging from mild skin infections to life-threatening diseases. One of the distinguishing features of S. aureus is its ability to produce an array of virulence factors, including toxins and enzymes, which contribute to its pathogenicity (1). The role of S. aureus in foodborne illnesses is of particular concern, where the production of enterotoxins can lead to intoxication rather than infection (2). Foodborne S. aureus intoxication is a significant public health issue, with outbreaks commonly associated with the consumption of contaminated foods such as dairy products, meat, and bakery items (3).
The regulation of virulence gene expression plays a pivotal role in its pathogenicity. Recent studies have suggested that natural compounds found in berries possess antimicrobial properties and may modulate the expression of virulence genes in bacterial pathogens (5,6). This study aims to explore the impact of berry compounds on the expression of virulence genes in S. aureus, focusing on the methodology developed by Nielsen et al 2010 (4). The S. aureus strains tested were isolates derived from NCTC8325 that are methicillin-susceptible according to established antimicrobial susceptibility profiles (4, 20).
The primary objective of this study is to explore berry extracts as selective modulators of S. aureus virulence genes, aiming for a nuanced control strategy that enhances food safety without complete bacterial elimination. The rationale behind this exploration stems from the need to identify novel strategies to combat S. aureus infections, informed by the documented antimicrobial properties of berries. The ability of S. aureus to produce toxins and enzymes presents a challenge in clinical and food safety settings. The toxins produced by this bacterium are heat-stable and resistant to many conventional food processing methods, making it difficult to eliminate them from contaminated food products (3). Ingestion of these toxins can lead to rapid-onset symptoms, including nausea, vomiting, and diarrhea, which can be severe and debilitating (3). Therefore, effective strategies to control S. aureus and its toxin production in food are of paramount importance for ensuring food safety and preventing outbreaks. In recent years, there has been growing interest in exploring natural compounds with antimicrobial properties as potential alternatives to conventional antimicrobials. Berries, in particular, have garnered attention due to their rich content of bioactive compounds, including polyphenols, flavonoids, and anthocyanins, which have been shown to possess antimicrobial activity against a range of pathogens (7). Previous studies have demonstrated the antibacterial effects of berry extracts against various foodborne pathogens, including Escherichia coli and Salmonella (7). However, the antibacterial potential impact of berries against S. aureus, specifically in the context of toxin production and gene expression, remains largely unexplored. This study aimed to investigate the influence of berry compounds on the expression of virulence genes in S. aureus, with a focus on their potential impact on toxin production. To the best of our knowledge, this is the first study to comprehensively examine the effects of berry extracts on the regulation of virulence gene expression in S. aureus. Understanding the interactions between berry compounds and S. aureus at the molecular level could provide insights into developing novel strategies to control toxin production in contaminated food and mitigate the associated health risks. The present study builds upon previous research that demonstrated berries' antimicrobial and preservative effects in food models (7). By investigating the specific effects of berry compounds on the expression of virulence genes in S. aureus, we aimed to shed light on their potential as natural antimicrobial agents that can target the underlying mechanisms of pathogenicity.
Bacterial Strains and Culture Conditions
The bacterial strains used are listed in and virulence gene expression.
| Table 1 Bacterial strains used in this study | ||||
| Strain | Agar | Broth | Source | |
| LacZ strains | ||||
| Staphylococcus aureus | TSA | TSB | 8325-4 -derived Hla: lacZ (PC322; Eryri) (4) | |
| Staphylococcus aureus | TSA | TSB | 8325-4 -derived palace (PC203; Eryri) (4) | |
| Staphylococcus aureus | TSA | TSB | SH101F7 radii: lacZ (4) | |
Table 1. The bacteria were cultured in nutrient-rich media to promote optimal growth Test for expression of virulence gene
The influence of berry compounds on the expression of virulence genes in S. aureus was determined by a method introduced by Nielsen et al 2010 (4). Briefly, overnight cultures of S. aureus 8325-4-derived strain PC322 hla::lacZ, PC203 spa::lacZ, or SH101F7 rnaIII::lacZ were prepared in TSB from single colonies grown on TSA plates. Two milliliters of 103 diluted overnight cultures were placed in a petri dish (12×12cm). Thereafter 50 ml of TSA (~40°C) containing 150 µg.ml-1 X-Gal and 5 µg.ml-1 erythromycin was added and mixed carefully. The plates were dried, and wells were cut with a sterile drill. Compounds were poured into the wells and plates were incubated at 37°C. The incubation period was until the appearance of a blue color (interval 13 to 48 h). The experiment was repeated with food ingredients added TSA to examine if the compounds have the same effect in food. The food ingredients included meat extract (10%, w/v), sunflower oil (5%, v/v), and starch (2%, w/v). Tested compounds included extracts from seven different berries (e.g. aronia, blackcurrant, blueberry, bilberry, cranberry, lingonberry, and raspberry) and purified anthocyanins from four of them (e.g. aronia, blackcurrant, blueberry, and cranberry). All the extracts have been neutralized and diluted in PBS as described earlier. Purified anthocyanins have been dissolved in absolute ethyl alcohol.
Plant Material and Berry Compounds
A diverse selection of berries, including Aronia (Aronia melanocarpa), blackcurrant (Ribes nigrum L.), blueberry (Ericaceae Vaccinium), cranberry (Vaccinium oxycoccus L.), and raspberry (Rosaceae Rubus) in dried powder form were purchased from Berrifine, Ringsted, Denmark. Bilberry (Vaccinium myrtillus L.), cranberry (Vaccinium oxycoccus & macrocarpon), and lingonberry (Vaccinium vitis-idaea L.) in spray-dried free-flowing extract form under the trade names of NutriPhy® Bilberry, NutriPhy® Cranberry and NutriPhy® Lingonberry were kindly provided by Chr. Hansen. These plants were selected based on their known bioactive components, such as polyphenols, flavonoids, and anthocyanins.
Extraction and preparation
Berry extracts were prepared using a standardized extraction protocol (7). Berry powders were extracted with 50% aqueous methanol or ethanol (100 mg×ml-1) (8,9). The mixture was shaken for 24 h at 40°C to ensure complete extraction (10). The extracts were filtered through Munktell G/3w paper under a vacuum, and the residue was repeatedly extracted with the same solvent until it was colorless (8). Subsequently, the extracts were passed through a 0.45 µm sterile filter (Syringe Filter Q-Max 25 mm 0.45 µm CA membrane sterile, Frisenette ApS). A portion of the extracts was taken for analysis of their composition and the rest was subjected to neutralization and evaporation of solvent or extraction of anthocyanins and purification.
Neutralization
The berry extracts were poured into a beaker and neutralized with sodium hydroxide (1M NaOH) under continuous stirring, pH was adjusted to 7 (PHM210 Standard pH Meter, MeterLab, France). The evaporation of neutralized extracts was performed at 40°C using a heater (RCT basic, IKAMAG) until dryness. Dried berry extracts were dissolved in phosphate-buffered saline (PBS). The extracts were collected in sterile screw cap tubes and kept at 4°C. Extraction of anthocyanins and purification
The crude extracts were further purified, by a packed bed adsorption method based on the work of (9) using AmberliteXAD-7column chromatography. A glass column (35×2cm) with a sintered glass bottom was filled with 50 ml of the resin (AmberliteXAD-7). The column was washed with a 4-bed volume of ethanol 99.9% and rinsed with a 4-bed volume of deionized water- sterile filtered (Syringe Filter Q-Max 25 mm 0.45 µm CA membrane sterile, Frisenette ApS). After the berries’ extracts were filtered, the resulting residue was subjected to the prepared column chromatography. The column was washed with 1 L of water, and elution of the anthocyanins was carried out with ethanol 99.9%. The eluate was concentrated at 40°C using the (RCT basic, IKAMAG) heater. Standardization of berry extracts concentration the volume of different berry extracts was determined based on their contents of 20 mg. ml-1 TAC, 20 mg. ml-1 TPhC, and 20 mg. ml-1 sum totals of TAC and TPhC.
Preparation of inocula
S. aureus NM was spread from glycerol stocks in TSA plates to get isolated colonies. After incubation of the plates overnight at 37°C, 3 to 4 colonies were taken and suspended in 5 ml of saline solution 1.9% (w/v) (FK). The solution turbidity was matched with the 0.5 Mac Farland standard (~106 colony forming units per ml) using a nephelometer (sensititre, nephelometer, Thermo). In this study, experiments were conducted with a single replicate to assess the impact of berry compounds on the expression of virulence genes in S. aureus. The use of a single replicate allowed for focused insights into the specific conditions tested, although it's acknowledged that replication is essential for broader generalizations.
The influence of seven different berry extracts on the expression of virulence genes of S. aureus was examined by the method introduced by (4). The method uses reporter strains with lacZ fused transcription to their central virulence genes and a chromogenic-galactosidase substrate. Compounds influencing the expression of virulence genes were noted based on color change on the plates. The plates were interpreted based on the appearance of blue color. High expression of the gene gives blue color whereas lack of blue (=white) shows down-regulation. All the results are presented in Fig 1, Fig. S1, Fig. S1, and Subfigure D illustrates the sequential arrangement of berry extract injections within individual wells on the agar plates. The media used in this study included TSA as the standard method (Fig. 1, column 1), TSA supplemented with starch (2%, Fig. 1, column 2), TSA supplemented with sunflower oil (5%, Fig. 1, column 3), and TSA supplemented with meat extract (10%, Fig. S1). Berry extracts could be categorized into three groups.
1. Neutralized berry extracts from Aronia (1st well of every plate), Blackcurrant (2nd well of every plate), Blueberry (3rd well of every plate), Cranberry (4th well of every plate), and Raspberry (9th well of every plate).
2. Purified anthocyanin content of berries, including Aronia (5th well of every plate), Blackcurrant (6th well of every plate), Blueberry (7th well of every plate), and Cranberry (8th well of every plate).
3. Concentrated solution of Nutriphy berries (including Lingonberry, Bilberry, and Cranberry, 10th, 11th, and 12th well of every plate, respectively) from Chr-Hansen (neutralized and in PBS).
All the neutralized berry extracts (1st group) reduced the expression of all three virulence genes, namely hla, spa, and RNAIII, in S. aureus on standard media, starch-supplemented media, and oil-supplemented media. Please refer to the subfigures of Fig. 1, rows a, b, and c, corresponding to well numbers one to four and nine. However, these effects could not be visualized on meat-supplemented media due to the intense color of the media, which is presented in the supplementary figure (Figure. S1).
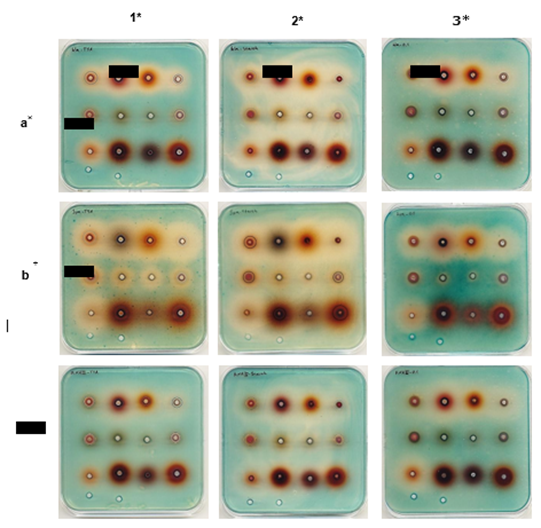
Figure 1: Influence of berry extracts on expression of three virulence genes (hla, spa and RNAIII)on TSA (standard assay)and TSA supplemented with starch 2% and oil 5%. Row a: Agar plates containingS. aureusPC322 (hla::lacZ); b: Agar plates containing S. aureusPC203 (spa::lacZ); c: Agar plates containing S. aureusSH101F7 (rnaIII::lacZ).
Column 1: TSA plates (the standard method); 2: Starch supplemented plates;
3: Oil- supplemented plates.
Fig. 1 Influence of berry extracts on expression of three virulence genes (hla, spa and RNAIII) on TSA
(standard assay) and TSA supplemented with starch 2% and oil 5%. Row a: Agar plates containingS. aureusPC322 (hla: lacZ); b: Agar plates containing S. aureusPC203 (spa: lacZ); c: Agar plates containing S. aureusSH101F7 (rnaIII: lacZ).
Column 1: TSA plates (the standard method);
2: Starch supplemented plates;
3: Oil- supplemented plates.
Order of extracts on the plates.
Aronia a′ Blackcurrant a′ Blueberry a′ Cranberry a′
Aronia b′ Blackcurrant b′ Blueberry b′ Cranberry b′ R
aspberry a′ Lingonberry c′ Bilberry c′ Cranberry c′
PBS Ethanol 99.9%
a′: Neutralized berry extract in PBS. b′: Purified berry anthocyanin in ethanol 99.9%. c′: Highly concentrated Nutriphy Berries from Chr-hansen (Neutralized and in PBS).
Nutriphy Berries are polyphenol normalized (to ≃30%) and spry dried berry extracts. * Inhibitory zones are clarified by black rings. Pictures are taken after 13 h (×) and 48 h (+) of incubation at 37°C.

Figure S2
Influence of berry extracts on expression of three virulence genes (hla, spa and RNAIII) on TSA (standard assay) and TSA supplemented with starch 2%, oil 5% and meat extract 10%.
Agar plates containing S. aureus SH101F7 (rnaIII::lacZ) Influence of berry extracts on expression of RNAIII
on TSA (a), TSA + 2% starch (b), TSA + 5% sunflower oil (c), TSA + 10% meat-extract (d). Order of extracts on the plates ►
a′. Neutralized berry extract in PBS
b′. Purified berry anthocyanin in ethanol 99.9%
c′. Highly concentrated Nutriphy Berries
Aronia a′ Blackcurrant a′ Blueberry a′ Cranberry a′
Aronia b′ Blackcurrant b′ Blueberry b′ Cranberry b′
Raspberry a′ Lingonberry c′ Bilberry c′ Cranberry c′
PBS Ethanol 99.9%
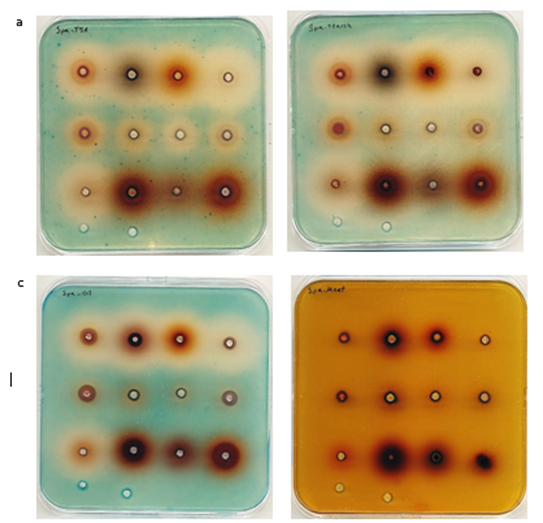
Figure S3
Influence of berry extracts on expression of three virulence genes (hla, spa and RNAIII) on TSA (standard assay) and TSA supplemented with starch 2%, oil 5% and meat extract 10%.
Agar plates containing S. aureus PC203 (spa: lacZ)
Influence of berry extracts on expression of spa on TSA (a), TSA + 2% starch (b), TSA + 5% sunflower oil (c), TSA + 10% meat-extract (d).
Order of extracts on the plates ►
a′. Neutralized berry extract in PBS
b′. Purified berry anthocyanin in ethanol 99.9%
c′. Highly concentrated Nutriphy Berries
Aronia a′ Blackcurrant a′ Blueberry a′ Cranberry a′ Aronia b′
Blackcurrant b′ Blueberry b′ Cranberry b′ Raspberry a′
Lingonberry c′ Bilberry c′ Cranberry c′
PBS Ethanol 99.9%
In general, the order of more influenced virulence genes by berry extracts (1st group) on S. aureus can be shown as spa > hla > RNAIII. The decreasing effects of berry extracts were at the lowest level in the expression of RNAIII, where here especially aronia and cranberry extracts had the lowest effect compared to other berries. The influence of purified anthocyanin content of berries (2nd group) was only observed in the reduction of spa [removed]i.e., Fig. 1 subfigures of row b). This influence was also more visible in standard and starch-supplemented media (i.e., Fig. 1 subfigures of row b, columns 1 and 2) compared to the oil-supplemented medium (i.e., Fig. 1, the last subfigure of row b). The effect of Nutriphy berries (3rd group) in this experiment was moderate based on their concentrations compared to the 1st group, as observed in the reduction of spa [removed]i.e. Fig. 1 subfigures of row b). However, the intense color of Nutriphy berries in the media posed a significant challenge for clear visualization. The production of various virulence factors by S. aureus is closely linked to the pathogenicity and virulence of this bacterium (11). These virulence factors encompass enterotoxins, toxic shock syndrome toxin-1 (TSST-1), cytolytic toxins (α, β, δ, and γ hemolysins), exfoliative toxins, Panton-Valentine leucocidin (PVL), cell-surface-associated proteins (such as protein A), and an array of enzymes (11,12). Among these virulence factors, alpha-hemolysin is described as the major lethal toxin of S. aureus [12]. Alpha homolysing is encoded by the hla gene (13,4). Bacterial cell adherent proteins such as protein A are described as fundamental factors for the pathogenesis of S. aureus infections, due to their facilitating role in binding of bacteria to host and biofilm formation on prostheses cardiovascular devices [14] Protein A is encoded by the spa gene [13.4]. Encoding several S. aureus virulence factors such as enterotoxins, δ-hemolysin, lipase, and exoproteases is dependent on the up-regulation of RNAIII [15].
The present study used standard and food – food-supplemented media to investigate the influence of berry extracts on the expression of three virulence genes (hla, spa, and RNAIII) in S. aureus in general and in the food composition. The berry extracts downregulated the expression of three virulence genes (hla, spa, and RNAIII) in the standard media and media supplemented with starch or sunflower oil.The influence of berry extracts on the expression of virulence genes was not observable in the plates containing meat extracts and Nutriphy berries, owing to the interference caused by the color of meat extract and the intensity of color exhibited by Nutriphy berries. However, the purified anthocyanin extracted from the berry samples exhibited a down-regulating effect, particularly noticeable in the case of S. aureus PC203 (spa: lacZ). It is noteworthy that even though the inhibitory zone was not specifically determined for raspberry extract, a discernible downregulation of virulence genes was evident (Figure. 1).
[13] performed a study on the effects of environmental factors on the expression of some S. aureus virulence genes such as hla, tst, spa, and RNAIII. They have investigated the effects of salt, sucrose, divalent cations (e.g. magnesium, calcium, and iron), and some chelating agents (e.g. EDTA, EGTA, and sodium citrate). They have observed that the presence of salt, sucrose, and divalent cations in the environment can have an interactive role in the control of S. aureus virulence determinants like hla, tst, spa, and RNAIII expression [13] Their results provide information that the presence of salt and sucrose can strongly repress the expression of hla, tst, and spa, whereas RNAIII is not altered. The addition of divalent cations can cause repression of tst, where, hla is not affected. Chelating agents can have an inhibitory influence on spa, not considerable on hla, and a simulative effect on tst [13]
The current study demonstrates that berry extracts effectively suppressed the expression of hla, spa, and RNAIII genes. Notably, the suppression effect was most significant on spa and least significant on RNAIII. It's recognized that within the accessory gene regulator (agr) system, the expression of RNAIII and spa exhibit a reverse relationship [16,15] Conversely, [13] reported spa and hla repression without altering RNAIII, suggesting the potential of independent regulation outside agr. Building upon the findings of [13] it could be hypothesized that the berry extracts' impact on spa and hla repression in this study is possibly potentiated by their sugar content. This notion is reinforced by the documented repressive influence of sucrose on hla, tst, and spa [13] Worth noting is that sucrose's suppression effect on hla and tst appears to be more pronounced than on spa [13] whereas our study observed a higher impact of berry extracts on spa. Moreover, the present study highlights that purified anthocyanins within berries also contribute to the repression of spa.[17] conducted a comprehensive review that highlighted the anti-adhesion properties of both berries and their phenolic extracts against various pathogens, such as Salmonella, E. coli, Helicobacter pylori, and S. aureus. The authors put forth the hypothesis that the inactivation and temporary immobilization of Salmonella, possibly induced by berries, might contribute to its anti-adhesion effect [17]. In addition to this, the inhibition of a gene responsible for encoding adhesion factors, such as the observed repression of spa in S. aureus, could be identified as an additional mechanism underlying the anti-adhesion effect of berries. The potential impact of berry extracts on tst expression warrants consideration, particularly given the presence of flavonoids known for their chelating properties [18] It's worth noting that chelating agents have been reported to stimulate tst expression, as established by [13] It's essential to clarify that the stimulating effect of chelating agents on tst expression is attributed to the leakage of cations in the media rather than the chelating agents themselves. This up-regulation of tst was effectively reversed by the excessive supplementation of divalent cations. Additionally, berries are acknowledged for their mineral content [19] and sucrose content. The complexity and diversity of compounds found in berries make addressing this question challenging and require further exploration. Overall, the suppressive influence of berry extracts on the expression of three virulence genes (hla, spa, and RNAIII) in S. aureus was observed in standard, starch, and oil-supplemented media. This influence was greater on spa and after that on hla and less on RNAIII. Several factors in berry extracts may be involved in this influence such as their sucrose, divalent cations, and anthocyanin content. A study by [6] delves into the antimicrobial potential of fruit extracts, including berries, against pathogenic bacteria. This investigation lends support to the concept of utilizing berry extracts to extend the shelf life of food items by curbing bacterial virulence and growth. In line with [6], this study underscores the potential reduction of food poisoning instances through the incorporation of berries or their derivatives. By curtailing the expression of virulence genes in S. aureus, these berry extracts hold promise in impeding the bacterium's capacity to incite infections and generate toxins—often culprits of foodborne illnesses.
The primary findings of this research spotlight the down-regulation of virulence genes through berry extracts, leading to a diminished manifestation of toxins and other virulence factors in S. aureus. The significance of S. aureus toxins in causing foodborne ailments has been addressed in other studies [3,1], further corroborating this research. Prior instances of inhibiting the growth of detrimental bacteria with berry extracts have been documented [7], underscoring the potential of employing natural compounds like berry extracts as additives for preserving food items. Such an approach offers a secure and natural means of curbing harmful bacteria growth and, consequently, minimizing the occurrence of foodborne illnesses.
This study investigated the impact of berry extracts on virulence gene expression in S. aureus. Distinct categories of berry extracts exhibited varying effects on virulence gene expression, with neutralized extracts showcasing a significant reduction in hla, spa, and RNAIII genes within S. aureus. Purified anthocyanin content and Nutriphy berry extracts also demonstrated down-regulating effects on specific virulence genes, albeit sometimes obscured by media color intensity. Notably, the neutralized extracts exhibited the most pronounced reduction in gene expression. The observed downregulation of virulence genes, especially spa, and hla, implies that berry extracts possess the potential to curb S. aureus pathogenicity by suppressing pivotal virulence factors. Moreover, these effects were consistent across diverse media compositions, underscoring the relevance for foodborne infection contexts and the plausible utilization of berries as natural antimicrobial agents. Significantly, the suppression of virulence genes correlates directly with lowered food poisoning risk. Reduced virulence factor production diminishes the likelihood of infections and associated symptoms upon consumption. This study suggests that integrating berry extracts, known for their capacity to down-regulate S. aureus virulence genes, could potentially enhance food product shelf life by thwarting pathogenicity and biofilm formation. Inhibiting virulence factors could curtail bacterial contamination and spoilage risks, thus enhancing overall food safety and preservation.
Critical Comment and Criticism
It is important to note that the specific mechanism by which the berry extracts exert their effects on gene expression was not fully explored in this study. Further investigation into the active compounds and their interactions with bacterial regulatory systems would provide a more comprehensive understanding of the observed effects.
Further research is needed to elucidate the specific compounds responsible for these effects and their mechanisms of action. Understanding the underlying molecular interactions would provide valuable insights into the potential development of targeted therapeutic interventions.
Potentials and Future Perspectives
Berries are found as more active against gram-positive bacteria (7), and they also decrease the expression of some virulence genes even in food compositions. Hence, the potential impact of berries on the growth and toxin production of S. aureus, B. cereus, and Clostridium species warrants exploration in future studies.
The initial experiment conducted on the purified anthocyanin content of berries and the concentrated solution of Nutriphy berries revealed a promising reduction in S. aureus virulence gene expression. Nevertheless, further experimentation and a deeper comprehension of the active compounds within berries are imperative for a comprehensive understanding of their potential effects.
The author extends their deepest gratitude to Professor Hanne Ingmer from the Department of Food Safety and Zoonoses at the University of Copenhagen for her financial support and contribution to this project, with special recognition to Dr. Jette Kjeldgård for her insightful idea of investigating the impact of berries on S. aureus virulence genes.
This study received funding through a master's thesis project conducted at the University of Copenhagen, overseen by Professor Hanne Ingmer (Food Safety and Zoonoses - Stigbojlen4, 1870 Frederiksberg C
The author (Massoud Attarianshandiz) affirms that there are no conflicts of interest to disclose.
Supplementary material
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
