
Research | DOI: https://doi.org/10.31579/2578-8949/099
1 Dermatology and Andrology department, Faculty of Medicine, Minia University, Egypt.
2 Pathology and Laboratory Medicine, London Health Sciences Centre, Schulich School of Medicine & Dentistry, Western University, London, Canada.
*Corresponding Author: Sherif S. Awad, Dermatology and Andrology department, Faculty of Medicine, Minia University, Egypt.
Citation: A Fabrice, A Hugues, D Juste, D Berenice, A Nadege, at el. (2022) Epidemiological profile of tumor dermatoses in the hospital setting in Cotonou (Benin) from 2009 to 2018. Dermatology and Dermatitis. 7(2); Doi:10.31579/2578-8949/099
Copyright: ©2022 Sherif S. Awad, this is an open-access article distributed under the terms of The Creative Commons. Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 26 May 2022 | Accepted: 25 April 2022 | Published: 09 May 2022
Keywords: vitiligo; immunomarkers; melanoblasts;melanocyte stem cells
Melanocytes are specialized neural crest-derived cells that are responsible for skin pigmentation. In vitiligo, loss of functioning epidermal melanocytes results in loss of pigment. Based on published studies, many transcription factors including microphtalmia-associated transcription factor, SRY (Sex Determining Region Y)-Box 10 are responsible for differentiation of melanocytes from neural crest cells as well as development of melanoblasts). However, the pathogenesis of loss of pigmentation in skin still unclear.
We studied the expression of several melanocyte-related proteins and markers in vitiligo samples and comparing them to normally pigmented skin samples. This may provide a more insight into the pathogenesis of vitiligo which is a still controversy.
We assessed the expression of six antibodies including S100, Melan-A (to detect fully formed, functioning melanocytes), CD117 (a specific relevant tyrosinase inducer), MITF and SOX-10 (important related transcription factors), and BCL-2 (stem cell markers) in 32 skin samples.
We demonstrated significant decline in the expression of all examined immunomarkers in the skin of vitiligo when compared to normal pigmented skin.
Our data may speculate that vitiligo pathogenesis does not involve only destruction of functioning active melanocytes, but also, other steps of cellular stimulation of induction of melanocytes and melanogensis are significantly affected as well.
microphtalmia-associated transcription factor (MITF), SRY (Sex Determining Region Y)-Box 10 (SOX10), tyrosine kinase receptor KIT (c-kit), melanocyte stem cells (MelSCs), Dopachrome tautomerase gene (Dct),
Melanocytes are specialized neural crest-derived cells that are responsible for skin pigmentation. The origin of melanocytes can be directly from neural crest cells migrating at the level of the skin through a dorsolateral migratory pathway, or from Schwann cell progenitors present in the peripheral nerves located at the level of the skin. Melanocytes reside in the basal layer of epidermis where they form the epidermal melanin units. The ratio of melanocytes to keratinocytes is 1: 10 in the epidermal basal layer (Haass et al. 2005). Melanocytes produce melanin in melanosomes and transfer it to neighboring keratinocytes by using the dendrites, giving the skin color, and protecting the genetic material of keratinocytes from damage caused by UV radiation (Pisarchik A, Wortsman J 2004).
Melanoblasts are the melanocyte precursor cells and arise during gastrulation of embryogenesis at the dorsal edge of the neural crest (Thomas AJ 2009). The differentiation of melanocytes from neural crest cells as well as development of melanoblasts is controlled through complex molecular mechanisms mediated by a network of transcription factors, including microphtalmia-associated transcription factor (Mitf), SRY (Sex Determining Region Y)-Box 10 (SOX10), Paired Box 3 and Wnt proteins (Hou, Ling & Pavan 2008).
A key factor during melanocyte differentiation is microphthalmia associated transcription factor (Mitf) (Widlund HR 2003), (Hemesath, Timothy & Steingrímsson, Eiríkur & Mcgill, Gaël & J Hansen, M & Vaught, J & Hodgkinson 1994). MITF controls the expression of key pigment synthetic genes including TRP-1, dopachrome tautomerase and tyrosinase (Lin, Connie et al 2002). Various extracellular signaling pathways converge on MITF to control both migration and survival of melanoblasts (Tabone-Eglinger S., et al 2012).
Dependency on tyrosine kinase receptor KIT (c-kit) receptor and its ligand stem cell factor (SCF) is well known (Murphy, M., Reid, K. et al 1992) ,(Cable, J., et al 1995) ,(Sarin, Kavita & E Artandi 2007). In the bulge region of the hair follicle, melanoblasts can differentiate into melanocytes or can remain as melanocyte stem cells (MelSCs) (Kubic D, Jennifer, et al 2008).
Sox10 is also a neural crest transcription factor crucial for specification, maturation, and maintenance of Schwann cells and melanocytes. One of the studies suggested that SOX10 binding sites contribute significantly to the cAMP responsiveness of the Mitf promoter (Huber, W.E., et al 2003). Some of in vitro and in vivo tests showed that some melanogenic genes are stimulated not only by Mitf but also directly by SOX10, a regulator of MITF, suggesting the existence of feed-forward loops between SOX10 and Mitf target genes. Without functional MITF, however, melanocytes can never mature and become pigmented, but when they remain unpigmented because they have mutations in the melanin biosynthetic genes, they stay alive and thrive (Bauer GL, et al 2009).
Melanocytes molecularly are recognizable by identification of melanocyte-specific proteins as TYR, TYRP1, TYRP2, melanosomal matrix proteins as Pmel17, MART-1, and transcription factors such as microphthalmia transcription factor (MITF) (Plonka PM, et al 2009).
In vitiligo, loss of functioning epidermal melanocytes results in loss of pigment and induction of leukoderma characterized by disfiguring macules and patches of depigmented skin.(Arican O and Kurutas EB 2008).
The aim of this work is to monitor the expression of several melanocyte related proteins in normal skin and to compare that to the expression in vitiligo skin. This may provide a more insight into the pathogenesis of vitiligo which is a still controversy.
Clinical data
The study included 32 skin samples obtained using 4mm punch probes. Half of them were obtained from center of depigmented vitiligo skin of non-segmental vitiligo cases and the other half from normally pigmented skin away from the lesions. Biopsies were immediately fixed in 10% formalin, routinely processed and embedded in paraffin blocks.
Immunohistochemical staining and scoring:
Immunohistochemical staining was performed on the paraffin-embedded specimens using a semi-automated Discovery autostainer (Dako EnVision FLEX+ detection system) and EnVision FLEX+ DAB+ Chromogen.
Tissues were exposed to 7 different primary antibodies including: 1-S100: polyclonal rabbit anti-S100 (code no.: IR504, Dako, North America) is used. Melanocytic cells were scored positive if they showed cytoplasmic or nuclear staining. 2- Melan-A: FLEX monoclonal Mouse Anti-Human Melan-A (code IR633, Dako, North America) is used. Cells were scored positive if they showed cytoplasmic staining. 3- CD117: polyclonal Rabbit Anti-Human CD117 (c-kit) antibody (code no.: A4502, Dako, North America) at a dilution 1:100 is used. Cells were scored positive if they showed cytoplasmic staining. 4- MITF (microphthalmia transcription factor): monoclonal Mouse Anti-Human MITF antibody (code no.: M3621, Dako, North America) at a dilution of 1:100 is used. Cells were scored positive if they showed nuclear staining. 5- SOX10 monoclonal Mouse Antibody SOX10 (code: SC-365692, Santa Cruz Biotechnology, Europe) at a dilution of 1:300 is used. Cells were scored positive if they showed cytoplasmic or nuclear staining. 6- BCL-2: monoclonal Mouse Anti-Human BCL-2 Oncoprotein (code no.: IR614, Dako, North America) is used. Cells were scored positive if they showed cytoplasmic or nuclear staining.
Slides scanning:
The slides were examined and scanned by using Aperio glass slide scanner at Department of Pathology and laboratory Medicine at Western University. The ScanScope scans microscope slides at 20X, 40X to a digital image that can be viewed with software that simulates the use of a light microscope. The svs files can be viewed and manipulated using a free software program called ImageScope. Representative photographs were captured using Aperio ScanScope™ technology at equivalent exposure times.
Immunohistochemical scoring:
Evaluation of the epidermal expression was carried by counting the number of positively stained cells within the epidermis in 1 mm length of stained tissue in the obtained digital slide-photos and multiplying the number by the intensity of positive staining; 1 for mild staining, 2 for moderate intensity and 3 for strong. Three independent doctors carried out the evaluation and median value for each case was recorded for further statistical evaluation
Statistical analysis
Clinical data were compared across the two groups using analysis of variance for differences in clinical parameters including age, sex, skin type, duration and activity of the disease and family history. A two-way repeated measures ANOVA with an interaction test and pairwise differences were used for comparing parametric quantitative data of two groups regarding the differences in the expression of six immunomarkers. All tests were two-sided, with a P-value of <0>
Clinical data
The present study included 16 patients with vitiligo vulgaris. Eleven female patients (68.8%) and 5 males (31.2%) were included in this study. The age of these patients ranged from 12 to 54 years old (mean 21 ± 10.9). The disease duration ranged from 1 to 20 years (6.81 ± 5.7). A positive family history of vitiligo presented in 4 patients (25%). Associated autoimmune diseases presented in 2 patients (12.5%) (one patient had alopecia areata, the other patient had idiopathic thrombocytopenic purpura). As regard skin type, 3 patients (18.8%) were skin type III, 10 patients (62.4%) were skin type IV and 3 patients (18.8%) were skin type V (Table 1).
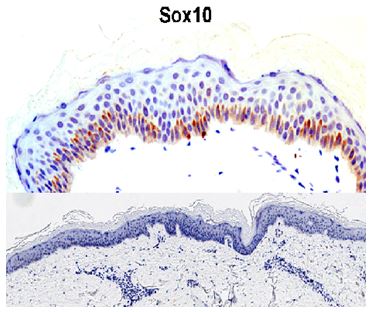
3.2 Results of immunohistochemistry staining:
S100 expression:
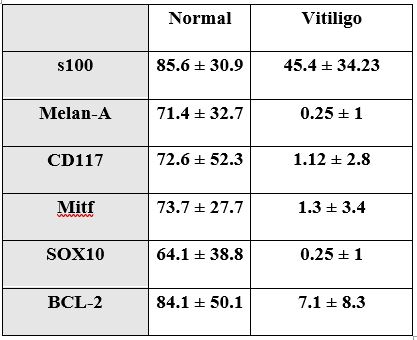
Examining the epidermis of the stained skin samples revealed that S100 positively stained cells were seen within the epidermis and at the basal layer. Scattered Cells were more abundant in normal skin compared to vitiligo skin which also contained several positively stained cells within the epidermis yet away from basal layer. S100 values in vitiliginous lesion ranged from 12 to 120 (mean 45.4 ± 34.23). In the non-lesional normally pigmented skin, it ranged from 45 to 144 (mean 85.6 ± 30.9) with statistically significant difference (p = 0.001), compared to vitiliginous lesion (Figure 1).

Melan-A expression:
Melan-A positively stained cells were demonstrated only within basal cells. Melan-A stained cells were minimal in vitiliginous lesion with values ranged from 0 to 4 (0.25 ± 1). In the non-lesional normally pigmented skin, it ranged from 15 to 135 (71.4 ± 32.7) with statistically significant difference (p = 0.001) (Figure 2).

CD-117 expression:
CD 117 (C-Kit) stained cells were demonstrated in the basal layer only. CD 117 stained cells in vitiliginous lesion were scanty and ranged from 0 to 10 (mean 1.12 ± 2.8). In the non-lesional normally pigmented skin, it ranged from 14 to 174 (mean 72.6 ± 52.3) with statistically significant difference (p = 0.001) (Figure 3).

MITF expression:
Mitf revealed positively stained cells within the basal layer. MITF stained cells showed values in vitiliginous lesion ranging from 0 to 12 (1.3 ± 3.4). In the non-lesional normally pigmented skin, it ranged from 7 to 108 (73.7 ± 27.7) with statistically significant difference (p = 0.001) (Figure 4)

SOX-10 expression:
SOX10 revealed positively stained cells within the basal layer. SOX10 stained cells in vitiliginous lesion were minimal with values ranged from 0 to 4 (0.25 ± 1). In the non-lesional normally pigmented skin, it ranged from 14 to 180 (64.1 ± 38.8) with statistically significant difference (p = 0.001) (Figure 5).
BCL-2 expression:
BCL-2 revealed positively stained cells within the basal layer. BCL-2 values in vitiliginous lesion ranged from 0 to 22 (7.1 ± 8.3). In the non-lesional normally pigmented skin, it ranged from 10 to 180 (84.1 ± 50.1) with statistically significant difference (p = 0.001) (Figure 6).

The overall expression values in vitiligo and normal skin are summarized in (Table 2 and Figure 7).

Correlation of expression of all markers in epidermis of diseased skin specimens:
By comparing the expression of all markers in the epidermis of vitiliginous specimens, significant decline was demonstrated. However, there is a significant difference between S100 & BC12 in expression in vitiligo specimens, but none of the others.
Melanocyte is a specialized cell which evolves from primitive skin stem cells and goes through a long and complicated process of differentiation, proliferation, migration, maturation and melanin synthesis. This study targeted the demonstration of expression of several melanocytes related proteins and markers in vitiligo samples and comparing them to normally pigmented skin.
S-100 and Melan A were used to detect fully formed and functioning melanocytes, CD117 was utilized as a specific relevant tyrosinase inducer, SOX10 and MITF were examined as important related transcription factors, and BCL-2 as stem cell markers.
It is known that, S100 protein can be expressed in different cells including fibroblasts, myoepithelial cells, histocytes and Langerhans cells in normal skin biopsies.(Xia J, et al 2016). We demonstrated that loss of S100 staining in vitiligo was mainly at basal layers as other non-melanocytic cells are still present intra-epidermally like Langerhans cells. Yet, significant drop in total values of expression in the epidermis was demonstrated due to the loss of functioning melanocytes located in the basal layer.
On the other hand, Melan-A expression is known to be restricted to melanin-producing cells, including normal and transformed skin melanocytes (Kawakami Y, et al 1994). Melan-A may be a melanosomal membrane protein, although its sequence does not bear any apparent homology to other known melanosomal proteins and does not possess melanogenic enzymatic activities(Kawakami Y, et al 1997). Our study was clearly demonstrating that Melan-A expression significantly decreased in vitiligo skin compared to normal skin. This data is comparable to previously reported studies and minimal persistence of some positive cells is also recorded before (Mandelcorn-Monson RL, et al 2003) (Park OJ, et al 2016).
CD117 was also examined as it is a known important and specific inducer of melanocytes and melanogenesis. Significant drop in expression of this was clearly demonstrated in vitiligo. The intracellular signaling from the c-Kit plays a critical role in the development of melanocytes. Downstream signal transduction molecules can be activated through the formation of the c-kit/SCF complex, and then regulate gene expression and cell growth, proliferation and differentiation (Nishikawa S, et al 1991), (Grabbe J, et al 1994), (Galli SJ, et al 1995). There was a significant decline in C-kit expression in vitiligo compared to normal skin and this decline is similar to previous reports although was non-significant in many cases (Mandelcorn-Monson RL, et al 2003),(Park O, et al 2016).
Transcription factors and proteins are required during the process of melanocytes induction and formation from precursors. The microphthalmia-associated transcription factor (Mitf) protein is a key regulator of melanocyte development. The function of Mitf has been particularly well-studied in melanocytes , and it has been shown that Mitf is involved in the maintenance and self-renewal of melanocyte stem cells (Lang D1, et al 2005) (Loercher AE, et al 2005). Mitf can efficiently transactivate the melanogenesis enzyme genes, such as tyrosinase and tyrosinase-related protein 1 (TRP-1) in cultured cells (Yasumoto K, et al 1994), (Yasumoto K, et al 1997) ,(Amae S, et al 1998) , (Shibahara S, et al 1999). A significant drop in expression of this factor was demonstrated in vitiligo samples compared to normal pigmented skin. Bhardwaj et al has been described similar findings in a recent study as well (Bhardwaj S, et al 2017).
SOX10 transcription factor belongs to the Sox-family of transcription factors important in the development and maintenance of melanocytes. It was significantly declined in vitiligo compared to expression in normal skin. In the melanocyte lineage, SOX10 directly regulates the expression of the Mitf gene (Bondurand N1, et al 2000), (Potterf SB, et al 2000), (Lee M, Goodall J, et al 2000) . and SOX10 also regulates the expression of the Dopachrome tautomerase gene (Dct), which functions in melanin biosynthesis (Britsch S, et al 2001) ,(Potterf SB, et al 2001) . It was demonstrated the elevation of expression of sox 10 in melanocytic tumors (Shin J, et al 2012) yet studying its expression in vitiligo is lacking and this could be first study to show its decline in vitiligo skin.
Stem cells are required to provide generations of melanocytes needed for the pigmentation process. No single marker is currently universally accepted as an identifier of all melanocyte stem cells (Bonchak JG, et al 2014) and here we examined for BCL-2. The proteins of the Bcl-2 family are important regulators of programmed cell death (Oltvai ZN, Milliman CL 1993) . BCL-2 interacts with Mitf to maintain survival of melanocyte (McGill GG, et al 2002). There is an indispensible role of BCL-2 in the development and maintenance of melanocyte stem cells (Hallsson JH, et al 2007) and so; mice null bcls lose pigment and melanoblasts decline on lose of BCL-2 (Mak SS, et al 2005). In our study BCL-2 was significantly decreased in the vitiliginous areas. Although some others, previously, denied any dysregulation of apoptosis molecules including BCL-2 in vitiligo (van den Wijngaar, et al 2000).
This study demonstrated the significant decline of several proteins and markers from the skin of vitiligo when compared to normal pigmented skin. It is comparable to many previous reports regarding MART-1, S100, and for a few reports regarding C-Kit expression or other transcription factors like Mitf yet it is the first report to signify the decline of SOX10, and BCL-2 in vitiligo skin.
In conclusion, Vitiligo skin lacks melanocytes and subsequently loses its melanocyte specific markers and proteins like Melan-A significantly. S100, although non-specific and can stain different types of cells within the epidermis, yet its expression in vitiligo also showed a significant drop compared to non lesional skin. The vital melanogenic factor C-Kit protein again demonstrated significant decline. Transcription factors and proteins required for induction of melanocyte and melanogenesis, Mitf and SOX10 were also significantly decreased. Stem cell related proteins and markers, BCL-2, significantly drop in vitiligo skin compared to non lesional skin and this may explain the persistent lack of the repigmentation in many cases and the characteristic recalcitrance of the disease.
Such findings may demonstrate that vitiligo pathogenesis does not involve only destruction of functioning active melanocytes, yet, other steps of early cellular stimulation of induction and melanocyte precursors are targeted and significantly affected as well.
This is a possible new view demonstrating that vitiligo could be a problem of homeostasis. In normal skin, not only functioning melanocyte proteins like Melan-A and CD117 exist but precursor and MelSCs related proteins like SOX-10, Mitf, BCL-2 also exist, so as to continually provide new generations of pigment producing cells to keep this pigmentary homeostasis. But in vitiligo, loss of those melanogenic machinery components leads to stoppage of production process from a very early step. Vitiligo skin cannot maintain the usual existence of melanocytes in the skin through continual stimulation of melanocyte precursor’s lineage due to lack of required precursors and their relevant proteins.
None
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
