
Research Article | DOI: https://doi.org/10.31579/2766-2314/090
1 BIO5 Institute, University of Arizona, 1657 E. Helen St, Tucson AZ 85718.
2 USDA Agricultural Research Service, Arid Land Agricultural Research Center, 416 W Congress St, Tucson AZ 85701.
3 Mycotoxin Prevention and Applied Microbiology Research Unit, USDA-Agricultural Research Service, National Center for Agricultural Utilization Research, 1815 N University St., Peoria, IL 61604.
*Corresponding Author: Monica A. Schmidt, BIO5 Institute, University of Arizona, 1657 E. Helen St, Tucson AZ 85718.
Citation: Monica A. Schmidt, Joseph Opoku, Hillary L. Mehl, (2023), Enzymatic Degradation of Aflatoxin Accumulation in Dry Maize Kernels as a Promising Post-Harvest Mitigation Strategy. J, Biotechnology and Bioprocessing 4(3); DOI: 10.31579/2766-2314/090
Copyright: © 2023, Monica A. Schmidt. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 26 April 2023 | Accepted: 11 May 2023 | Published: 16 May 2023
Keywords: maize; aflatoxin; enzyme; degradation; food security; post-harvest
Background: Aflatoxin is one of the most problematic fungal-produced toxins as it is responsible for massive global agricultural losses and is deleterious to both human and animal health. Contamination of crops by certain strains of Aspergillus fungi accumulate aflatoxin in both post- and pre-harvest conditions.
Methods: In this report, we tested the aflatoxin-degradation efficiency of an endogenously expressed enzyme in harvested maize kernels. In post-harvest conditions, equivalent loads of A. flavus were used to infect harvested maize kernels previously engineered to express an aflatoxin-degrading enzyme from the Honey mushroom fungus.
Results: No measurable, or significantly reduced, levels of aflatoxin were detected in all the enzyme-expressing harvested kernels initially and then 3 days post –harvest the transgenic kernels amassed aflatoxin.
Conclusions: This is the first report of an enzyme degradation of aflatoxin in a crop in harvested kernels. This demonstrates the potential of this strategy to aid in the mitigation of aflatoxin in post-harvest conditions.
Mycotoxins are toxic fungal-produced compounds that cause global economic and health issues. Estimates currently cite as much as 80percentage of world crops are contaminated with detectable levels of one or more mycotoxins [1] with at least 25percentage of crops exceeding regulated Codex Alimentarius limits [2]. Although there is difficulty in accurately measuring the prevalence of mycotoxins in global crops, the consumption of these compounds in diets has been correlated to deleterious health effects in humans and animals. These fungal-produced secondary compounds are responsible for massive economic losses and they adversely affect global trade. Aflatoxins are considered the most toxic group of mycotoxins as they are known carcinogenic compounds, being linked to liver cancer [3-4], implicated in the stunting of children’s growth [5] and sometimes cause acute death [6]. Aflatoxin-producing Aspergillus species can infect many crops but have an affinity for cereals and nuts. Maize is often the most impacted crop, with annual global losses due to aflatoxin contamination of $160 million in the US [7] and $450 million in sub-Saharan Africa [8].
Infection of maize with aflatoxin-producing Aspergillus species, including A. flavus, can occur both pre-harvest while the crop is actively growing and in post-harvest during storage. Pre-harvest aflatoxin mitigation strategies include breeding [9] or genetic engineering [10-13] for fungal resistant cultivars, use of RNAi suppression technology to inhibit aflatoxin production [14-16], competitive exclusion of aflatoxin-producing fungi using non-toxigenic strains [17-18], and agricultural practices such as insect control and fungicide applications. Post-harvest strategies largely focus on proper storage with moisture levels of particular concern as Aspergillus species grow in moist, humid environments, insect control as bite wounds serve as a gateway for the opportunistic fungus, low oxygen or high ozone environments and detection and sorting of contaminated kernels/cobs ([19] for review of post-harvest mechanisms).
A promising post-harvest decontamination strategy involves the enzymatic conversation of aflatoxin to innocuous compounds. Prior research [20] demonstrated the successful pre-harvest degradation of aflatoxin from contaminating A. flavus conditions in developing maize kernels that were expressing an endoplasmic reticulum (ER)-targeted embryo-expressed aflatoxin-degrading enzyme isolated from the edible Honey mushroom (Armillariella tabescens) [21]. In this research, we investigate if reducing /eliminating aflatoxin by enzymatic degradation in maize kernels would also be an effective post-harvest strategy in dry stored kernels.
Plant Material
Dry kernels were harvested from a control nontransgenic maize plant (Null) and three independent transgenic maize lines (Enz7, Enz8, Enz10) (Zea mays Hi II hybrid A 188 and B73 background) previously shown to be expressing an inserted aflatoxin-degrading enzyme (GenbankAccession AY941095) [20]. The inserted cassette in these transgenic maize plants consisted of a codon-optimized 2.166 kb open reading frame of an ER-targeted aflatoxin-degrading enzyme expressed by an embryo-specific maize globulin-1 promoter (Genbank Accession AH001354.2) [20]. Cobs from three transgenic lines (Enz 7, Enz 8, and Enz 10) and the nontransgenic maize line (Null) were removed from ears grown in the greenhouse. Kernels from the 3 transgenic enzyme-degrading aflatoxin and null control maize plants were harvested, dried, and stored at room temperature until they were used for infection experiments.
Aspergillus flavus inoculum preparation
Aspergillus flavus isolate AF13 [22] was obtained from the USDA-ARS Aflatoxin Biocontrol Lab culture collection. A granule from a silica gel stock vial of the isolate was transferred to 5/2 agar (5percentage V-8 vegetable juice, 2percentage agar, pH 5.2) and the plate was incubated in the dark at 31°C for 5 days. Approximately 10 agar plugs colonized with mycelia and spores of the isolate were transferred to a vial containing 3.5 ml of sterile water, and this was used as the working stock for growing up inoculum. Conidial suspensions (15 µl) from the water vial stock were transferred to wells in the center of 5/2 agar plates. After incubation at 31°C for 6 days, conidia were picked up from plates using sterile cotton swabs and suspended in 10 ml of sterile 0.02percentage Tween-80. Conidial suspensions were vortexed and transferred into nephelometric turbidity unit (NTU) vials, and turbidity was measured using a calibrated turbidimeter (model 965-10, Orbeco-Hellige, Farmingdale, NY) as described previously [23]. The final spore concentration was calculated using a standard curve for NTU versus spores/ml using the formula: spores/ml = NTU × 49,937. The spore suspension was diluted to a final concentration of 50,000 spores/ml in sterile water.
Kernel inoculation
Five-gram samples of kernels from each maize line were disinfested by submerging kernels in 10percentage commercial bleach (sodium hypochlorite) for 3 min. followed by 70percentage ethanol for 5 min. and a final rinse in sterile water. Kernels were air dried on sterile paper towels in a biosafety cabinet then transferred to 250 ml Erlenmeyer flasks (5 g per flask). The moisture content of a subsample of each maize line (4 g) was measured using a Halogen Moisture analyzer (Model HC103/03, Mettler Toledo, Columbus, OH). The moisture content of kernels in each flask was then adjusted by adding 10 µl (5 × 104 spores) of A. flavus isolate AF13 in the volume of water needed to bring kernel moisture to 25percentage. Flasks were then gently shaken to evenly coat kernels with the inoculum. For the non-inoculated control of each maize line, only sterile water was added to flasks. Flasks were sealed with gas permeable Bug Stopper plugs (Whatman, Piscataway, Nj) and transferred to an incubator set at 31°C. Three replicate flasks of each maize line were taken out of the incubator one, two, and three days after inoculation. Spores were washed off the kernels by adding 20 ml of 0.01percentage Tween-80 to each flask followed by shaking at 150 rpm for 10 min. on a Model HS 501 Digital Shaker (IKA Works Inc., Wilmington, NC). Spore suspensions were filtered through Miracloth and collected in 50 mL Falcon tubes. Kernels were washed again with 20 ml of sterile water, and the suspension was again filtered through Miracloth into the 50 mL tube. The ~40 mL spore suspensions were vortexed, and spore concentrations were measured using turbidity as described above. Washed kernels were dried at 60 °C for 36 h then ground for 35 s using an IKA A11 basic S1 grinder (IKA Works Inc., Wilmington, NC).
Aflatoxin quantification
For each sample, total aflatoxins (aflatoxin B1 + aflatoxin B2) were extracted from 1.5 g ground kernels with 15 ml of 70percentage methanol. Extracts were separated using thin layer chromatography (TLC) and aflatoxin was quantified using scanning densitometry as described previously [20, 24]. Briefly, 12 µl of extract was spotted on 20 × 20–cm TLC glass plates (Silica Gel 60 F254, Millipore) along with an aflatoxin standard (Aflatoxin Mix Kit-M, Supelco, Bellefonte, PA), and plates were developed with diethyl ether: methanol: water (96:3:1). The presence or absence of aflatoxins B1 and B2 were confirmed visually under ultraviolet light (365 nm) and quantified on plates using scanning fluorescence densitometry with a CAMAG TLC Scanner 3 (Camag Scientific Inc., Wilmington, NC). Quantities of aflatoxin relative to the standard were used to calculate total nanogram (ng) aflatoxin per gram (g) kernels (equivalent to parts per billion; ppb).
Means and standard error for all metrics measured from the maize kernel transgenic lines were determined. All quantitative metrics were performed in triplicate. Significant difference from the nontransgenic control to any experimental line was determined by student T-test analysis, plessthan0.05 levels.
An aflatoxin-degrading enzyme from the Honey mushroom Armillariella tabescens was subcellularly targeted to the ER to enhance its stability in maize kernels. The localization of inserted enzymes into the endomembrane system is a technique that has been demonstrated to be effective at both enhancing an enzyme’s stability and allowing an introduced enzyme’s accumulation. The ER-targeted enzyme cassette was under the regulation of an embryo-specific promoter as previous research had shown embryo metabolic activity was positively correlated with aflatoxin resistance in maize cultivars [26]. The inserted aflatoxin-degrading enzyme was detected via mass spectroscopy analysis of ground transgenic kernel tissue in all three transgenic Enz lines in both developing kernels and dry stored kernels [20]. In pre-harvest conditions, previous research on ER-targeted aflatoxin-degrading enzyme expressing maize kernels found undetectable aflatoxin accumulation in all 3 transgenic Enz maize lines after 14-days of infection and significantly reduced (90percentage reduction) aflatoxin loads after 30-days, compared to nontransgenic control levels [20]. To determine the effectiveness of this successful pre-harvest strategy in a post-harvest situation, non-inoculated aflatoxin-degrading Enz transgenic maize kernels were harvested, dried, and stored at room temperature prior to being infected with A. flavus strain AF13.
Inoculated kernels were incubated for 1 day, 2 days or 3 days at which time the aflatoxin load was quantified by TLC analysis. Kernels were incubated under non-optimum conditions for storage (e.g., warm temperature and high moisture/humidity), but conditions simulated those that could occur in poorly maintained storage environments. After 1-day, infected dry null kernels had an average 2.5 +/- 0.3 ppb log aflatoxin load compared to undetectable aflatoxin amounts in all three Enz transgenic maize kernels (Figure 1).
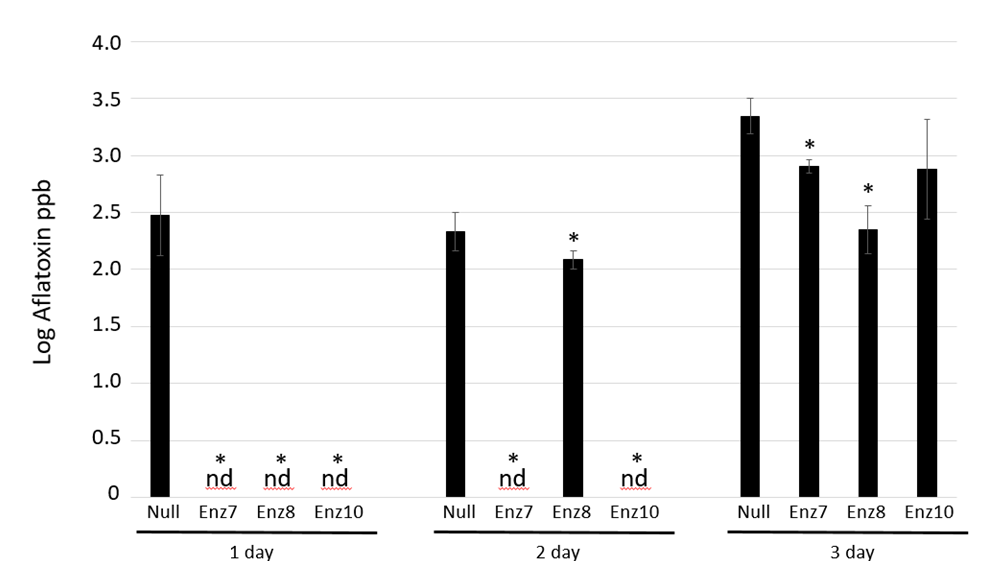
Harvested kernels were inoculated with Aspergillus flavus strain AF13 and infection was allowed to occur for 1 day, 2 days or 3 days. Aflatoxin loads were quantitated via thin layer chromatography (TLC) analysis at the three time points. Log aflatoxin means ± SE are shown. * Denotes significant difference from the nontransgenic control at plessthan0.05 levels, based on student T-test. Replicates were performed in triplicate. nd denotes not detectable at a limit of 20
Figure 1: Harvested Kernel Infection Assay of Aflatoxin-degrading Transgenic Maize
After 2 days of infection, the null kernels had 2.3 +/- 0.1 ppb log aflatoxin compared to undetectable in two Enz transgenic lines (Enz7 and Enz10) and 2.1 +/- 0.1 ppb log in Enz 8 line. The TLC methodology used to quantitate aflatoxin has a detection limit of 20 pbb (log value 1.3 ppb), hence the dry kernels with non-detectable aflatoxin loads had less than 20 pbb. Three days post-Aspergillus inoculation, null kernels contained 3.3 +/- 0.2 ppb log compared to Enz7, Enz8 and Enz10 having 2.9 +/- 0.1 ppb log, 2.3 +/- 0.2 ppb log and 2.9 +/- 0.4 ppb log aflatoxin, respectively. After a single day after inoculation, kernels from all 3 transgenic aflatoxin-degrading maize lines had no detectable aflatoxin accumulation compared to considerable levels already accumulated in the null controls. At two days post Aspergillus-infection, two (Enz7 and Enz10) of the three transgenic maize lines still displayed undetectable amounts of aflatoxin while one line (Enz8) had some aflatoxin accumulation but was significantly lower than the amount accumulated in the null controls as determined by student t-test plessthan0.05. Three days after inoculation, all dry kernels, both transgenic and null, accumulated aflatoxin with two of the lines (Enz7 and Enz8) accumulating significantly lower amounts compared to null controls (student t-test plessthan0.05). Considering most countries have regulatory aflatoxin limits around 20 pbb of aflatoxin as a consumption threshold, this methodology would be able to keep maize kernels within an acceptable range in short-term post-harvest conditions.
To investigate if the Enz transgenic maize dry kernels had reduced aflatoxin accumulation because they had a reduced initial Aspergillus infection, we quantified the amount of fungal growth on each sample by spore counts. Figure 2 shows the results of fungal spores detected in all samples across the 3 days of the infection experiments. After 1 day infection, spores on the null kernels were 7.07 +/- 0.2 log spore count compared to Enz7, Enz8 and Enz10 having 6.65 +/- 0.2 spores, 6.78 +/- 0.3 log and 6.50 +/- 0.1 log, respectively. Day two post infection, no significant difference in spore count was noted as null kernels had 7.84 +/- 0.1 log, compared to the three transgenic maize lines Enz7, Enz8 and Enz10 had 7.43 +/- 0.1 log, 7.40 +/- 0.3 log, 7.60 +/- 0.3 log, respectively. Finally, day 3 post infection, spore count on null kernels was 7.61 +/- 0.1 log compared to Enz7, Enz8 and Enz10 being 7.66 +/- 0.02 log, 7.70 +/- 0.01 log and 7.61 +/- 0.04 log, respectively. Consistently, no Enz transgenic dry kernels samples were determined to be significantly different from the null controls regarding the number of spores detected. This indicates that the Aspergillus infections were done consistently, and the amount of fungal inoculum and subsequent growth were comparative across all samples, yet when correlated with aflatoxin accumulation (Figure 1) it demonstrated the inserted aflatoxin-degrading enzyme was, at least initially, able to degrade the produced aflatoxin from the contaminating A. flavus.
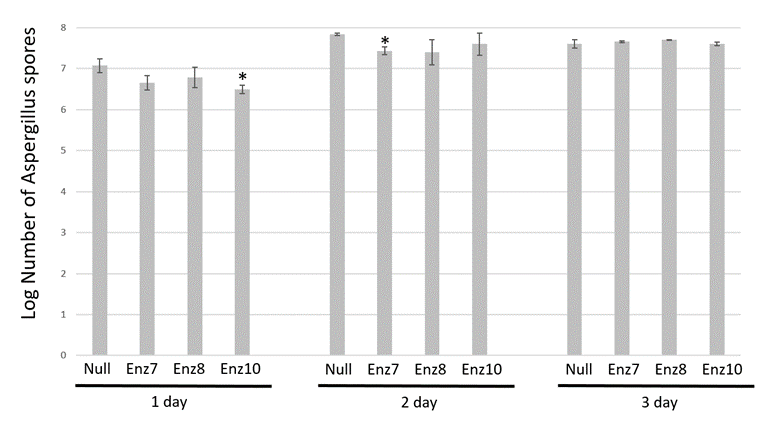
Spore counts are shown as log mean ± SE of three replicates. * Denotes means are significantly different from null nontransgenic as determined by student T-test analysis, plessthan0.05.
Figure 2: Spore count comparison of Aspergillus-infected transgenic maize harvested kernels.
Enzymatic degradation has been an effective post-harvest aflatoxin mitigation strategy typically employed by the exogenous mixing of contaminated food items with a microorganism, or its isolated degradation enzyme, capable of converting aflatoxins to innocuous compounds [25, 27].
This research is the first report of engineered crop plants capable of degrading contaminating aflatoxin in post-harvest conditions. Future reiterations of this research could focus on the enhancement of the aflatoxin-degrading enzyme accumulation in seeds, perhaps by expressing the enzyme under a strong kernel-specific promoter.
Aflatoxin has been a major global economic and health concern for decades and it is predicted that this food contaminant will increase in both frequency and severity if global climate temperatures continue to increase [28]. The degradation of accumulated aflatoxin by contaminating Aspergillus species on food items by engineered crops might play a role in eliminating, or at least reducing, this carcinogenic compound thereby enhancing global food security and safety.
This work was financially supported by United States Department of Agriculture National Institute of Food and Agriculture from the Improving Food Safety program, grant # 2019-67017-29644, title “Characterization and Enhancement of Higs in Aflatoxin-Free Transgenic Maize”. We thank Cyrus Mao for maintaining and harvesting the transgenic Enz maize kernels. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer.
The authors declare they have no known competing financial interests that might influence the work reported in this paper.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
