
Research Article | DOI: https://doi.org/10.31579/2690-1919/241
1 Centre for Human Reproduction 721, G.T.B. Nagar, Jalandhar-144001, Punjab, India.
2 Ex-Rotunda-A Centre for Human Reproduction 672, Kalpak Garden, Perry Cross Road, Near Otter’s Club, Bandra(W)-400040, Mumbai, India.
3 Consultant Neurologist, Swami Satyanand Hospital, Near Nawi Kachehri, Baradri, Ladowali road, Jalandhar, Punjab, India.
*Corresponding Author: Kulvinder Kochar Kaur, Centre for Human Reproduction 721, G.T.B. Nagar, Jalandhar-144001, Punjab, India.
Citation: Kulvinder K Kaur, Gautam Allahbadia and Mandeep Singh. (2022). Association of Iron Metabolism Abnormalities as Etiopathogenetic Factor in Alteration of Beta Cell Function and Impairment in Generation of Diabetes Mellitus: A Systematic Review. J Clinical Research and Reports, 11(1); DOI:10.31579/2690-1919/241
Copyright: ©2022, Kulvinder Kochar Kaur. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 10 March 2022 | Accepted: 25 March 2022 | Published: 04 April 2022
Keywords: ironmetabolism; β cells function; ROS; DM; Alzheimer’s disease; DNA; IR
Iron constitutes an essential element that is implicated innumerous physiological functions. In the context of the pancreatic β cells, they act as components of the Fe –S cluster proteins, which are a must for the appropriate insulin generation and processing. As far as mitochondria are concerned , in the form of a constituent of the respiratory chain it aids in the generation of ATP along with Reactive oxygen species(ROS),that result in induction of β cells depolarization that causes potentiation of insulin liberation that is calcium based .It is of great importance that a marked fiine tuning gets established with regards to the iron cellular amounts to guarantee normal provision of Iron along with avoidance of iron overload. Actually, in view of the great reaction with oxygen in addition to the generation of free radicals , iron overload might result in Oxidative injury of cells that possess susceptibility to this situations in view of the normal escalation of ROS development besides lesser availability of antioxidant enzymes action .Thus here we conducted a systematic review utilizing usual search engine utilizing the MeSH; iron metabolism; DM; haemochromatosis; thallasemia; Alzheimer’s ;Parkinson’s disease ; Friedrich’s ataxia; Iron homeostasis; Iron binding protein; transferrin bound iron(TBI); non TBINTBI); Divalentmetal transporter I(DMT1); ferroportin; islet amyloid polypeptide; zinc transporter ZIP 14; Chaperone proteins- poly CR binding proteins(PCBPs); mitoferrin(Mfrn); Fe-S clusters - enzyme CDKAL1; hepcidin; hephaestin; frataxin ; labile iron pool (LIP); ABCT7; PDX1;MafA; PHD; MAMs; Miner 1;gestational DM; ferroptosis; ferroportin; iron overload &treatment ;toxicity in brain, GIT; from 1980 till 2022 till date. We found a total of 4500 articles out of which we selected 135 articles for this review. No meta-analysis was done. Main aim of this review was to get a better insight in mode of iron homeostasis in β cells, with mode of changed in this event in their damage. How abnormal iron storage/chaperon proteins might cause diabetes.
Iron has a main part in numerous cellular functions like oxygen transportation along with exchange, constituting the metal part that possesses the capacity of development of Reactive oxygen species (ROS) which has the ability of injuring the DNA, phospholipids along with proteins. Hence it holds considerable significance with regards to cells in addition to organisms for the sustenance of homeostasis which sees to it that provision of Iron besides accrual of escalated Iron. Indeed, numerous disease events states are resultant of abnormal tackling of iron. Aberrations of iron homeostasis has been the properties of abnormal tackling of iron. This has been the observation in haemochromatosis, anemia, atherosclerosis along with diseases that implicate the central nervous system (CNS) like Alzheimer’s disease as well as Parkinson’s diseases, Huntington’s, Friedrich’s ataxia in addition to eating disorders like pica [1-5].
Escalation of proof has suggested an etiological part of iron in type2 diabetes mellitus (T2DM). Iron is necessary for insulin liberation [6], however, its collection is of significance in the causation of pancreatic islets inflammation, besides being believed to be a biomarker of risk of diabetes along with mortality [7].
The association amongst iron along with diabetes was initially picked up in case of patients of haemochromatosis in addition to β thallasemia [8,9], where abundance of iron was implicated in failure of β cells as well as insulin resistance (IR) was emphasized.
Additionally, in T2DM patients’ escalation of amounts of ferritin [10], that is a biomarker of enhancement of body iron stores as well as reduction of hepcidin amounts, that is the hepatic hormone implicated in systemic iron homeostasis have been the observation in blood, that emphasizes the systemic changes in iron metabolism [11].
Inspite of the collection of proof, the insight in the context of molecular modes that are correlated with escalation of iron as well as diabetes is absent. Moreover, whereas its homeostasis might have influence on separate organs, the islets of Langerhans appear to have specific proneness to iron. The collection of ROS taking place in the, existence of enhancement of iron possesses the capacity of resulting in Oxidative injury of the pancreatic β cells. The robustness of vulnerability towards Oxidative stress (OS) of these cells is secondary to enhancement of metabolic action [12,13], in addition to escalated generation of ROS in combination with the lowered defense modes against Oxidative insults. Actually, β cells possess the properties of reduction in expression of super oxide dismutase (SOD), catalase (CAT), as well as glutathione peroxidase (GPx) [12,14]. With the knowledge of escalation of failure of β cells in the generation along with propagation of diabetes here our Objective is to detail the progression with regards to tackling of iron along with iron cross talking proteins in β cells physiology in addition to pathology. A greater insight of iron metabolism along with homeostasis in these particular cells might aid in the generation of innovative treatments for the management of diabetes mellitus .Earlier we had reviewed lot of articles with regards to etiopathogenesis that was inclusive of gut microbiota, hypothalamic gliosis, macrophage polarization alterations, genetic predisposition, obesity, epigenetic alterations in obesity and treatment Diabetes mellitus[(DM)(both type1&type2)] [15-34]besides lot of novel modes of treatment. Here we attempt to correlate another etiological factor with the influence of iron metabolism.
Here we conducted a systematic review utilizing search engine PubMed, google scholar ;web of science ; embase; Cochrane review library utilizing the MeSH terms like the generation of DM, namely iron metabolism; DM; haemochromatosis i; β thallasemia; Alzheimer’s disease; Parkinson’s diseases; Friedrich’s ataxia; Iron homeostasis; Iron binding protein; transferrin bound iron(TBI); non transferrin bound iron(NTBI); iron regulatory proteins (IRPs); Divalent metal transporter I(DMT1); ferroportin; islet amyloid polypeptide; zinc transporter ZIP 14; Chaperone proteins- poly CR binding proteins(PCBPs); mitochondrial iron transporters mitoferrin (Mfrn); Fe-S clusters - enzyme CDKAL1; electron transport proteins; hepcidin; hephaestin; frataxin; labile iron pool (LIP); ABCT7; PDX1; MafA; prolyl and asparaginyl hydroxylase (PHD); Mitochondria correlated ER (endoplasmic reticulum) membranes (MAMs); Miner 1; gestational DM; ferroptosis; ferroportin; iron overload & treatment; toxicity in brain, GIT; from 1980 till 2022 till date.
We found a total of 4500 articles out of which we selected 135 articles for this review. No meta-analysis was done.
2. Homeostasis of Iron in β cells
In view of its chemical feature in addition to probable deleterious actions, cells have generated a complicated system in the context of tackling Iron: transportation of the ion by both carriers in addition to receptor subsequent to binding across the membranes, enzymes along with buffering proteins regulate the redox status along with free amounts, besides iron binding proteins, as per the ion amount. There is existence of numerous of these proteins in the pancreatic β cells, despite us not getting insight with regards to certain particular actors of this applicable event till date (figure 1) [revin 35].
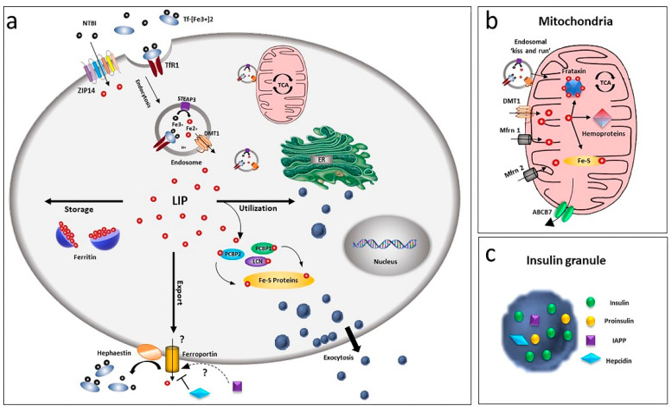
2.1 Influx of Iron via the plasma membrane
Iron uptake in β cells occur by 2 separate systems i) a receptor modulated transportation for the transferrin bound iron (TBI) as well as a non transferrin bound iron (NTBI) shift. The initial mode is dependent on the crosstalk of (TBI with the particular cell surface transferrin receptor 1 (TfRI) [6,36]. This complex subsequently gets internalized in the endocytic partition with the Divalent metal transporter I (DMT1 or SLC11A2) as well as the metalloreductase six transmembrane epithelial antigen of the prostate (STEAP3) [37].
The vesicles subsequent to internalization undergo fusion with the lysosomal chambers along with the acidic environment causes stimulation of the conformational alterations of Tf-Fe –complex in addition to the liberation of Fe 3+, that aids in the reduction of ferrous form through STEAP3. Expulsion of this Fe 3+ occurs in the cytoplasm via DMT1, that makes use of the H+ gradient that gets generated by the vacuolar H+ -ATPase (v- ATPase) in the form of the pushing force [38].
More recently, a NTBI uptake has further been the observation in the human beta cell line βlox5 [39].The precise chemical entity of plasma NTBI is not clear however it is thought to be existent basically in ferric citrate along with other low molecular weight species [40]. In certain pathological situations, higher molecular weight NTBI plasma fractions has been the observation, that pointed that probably binding of Fe 2+ as well as Fe 3+ to the proteins [41] in addition to the presence of separate NTBI pools, that is based on the iron overload situations [42]. NTBI might get found in the blood of patients in possession of iron overload situations on saturation of transferrin amounts [43], despite its existence has been the observation when saturation of transferrin amounts is not total [44]. Intriguingly in patients with diabetes NTBI existence is there at the saturation of transferrin under 60% [45]. In case of primary human islets of langerhans, modulation of NTBI occurs by the zinc transporter ZIP 14 (SLC39A14) that resides in the plasma membrane of the β cells at a place where iron overload is limited. Chronically high glucose amounts over24h results in up regulation of the expression of the transporter hence corroboration of the functional significance of ZIP 14 as well as pointed the probable sequelae of iron homeostasis [45]. Nevertheless, si RNA modulated ZIP 14 knocks down decided just by 50percentage decrease of NTBI uptake, pointed towards the implication of other transport systems. A part of L or T kind calcium channels appears not probable in view of absence of iron overload in murine β cells that display their expression [46].
2.2 Efflux of Iron via the plasma membrane
How Iron leaves the β cells remains in controversy; ferroportin/Ireg 1(FPN1, SLC40A1)thus far remains the only exporter for iron believed to exist in our knowledge[47], as well as islets demonstrate markedly low immunoreactivity with regards to this transporter [48] despite expression of hephaestin by them. This specific protein is implicated for the stabilization of the ferroportin, besides induction of its internalization [49], hence pointed to a positive feedback mode for Iron control at the time of glucose stimulated insulin secretion (GSIS) that gets modulated by ferroportin regulation [50].
An alternative probable ferroportin modulator is the islet amyloid polypeptide (IAPP) [51], that gets liberated along with insulin playing a part in glucose homeostasis [52], besides in regulation of food consumption [53]. Despite, its part in Iron homeostasis in β cells has not got proved, it might point to correspondent role in neurons, where amyloid polypeptide (APP result in stabilization of ferroportin at the plasma membrane in addition to cause induction of liberation of Iron via ferroxidase action [54-56], hence avoidance of iron overload besides Oxidative stress (OS).
2.3 Proteins that are Iron binding
With a good regulation of Iron homeostasis, cells prevent enhancement of deteriorating free iron. On gaining entry in the cells, they generate a cytoplasmic labile iron pool (LIP)which gets concealed by ferritin, that is the only cytosolic iron storage protein. Expression of H as well as L chains, both occurs in β cells, besides modulation at the translational level by iron overload; once there is escalation of iron, escalation of ferritin generation in addition to storage of ferritin occurs [57]. Via cloistering of this element, ferritin has a significant part in detoxification of iron, besides working in the form of iron reserve protein. Despite, the existence of a cytoplasmic LIP that comprises of iron that can be chelated has got observed earlier, there have been botheration with regards to iron on internalization in the cells gets transported to ferritin through direct protein- protein crosstalk in a hydrophobic milieu, as LIP does not appear to possess the properties of an intermediate iron pool [58]. Chaperone proteins, like poly CR binding proteins (PCBPs) [59], get implicated in this event. All the 4 PCBP isoforms whose knowledge we possess have the capacity of binding as well as transportation to cytosolic ferritin [57], however displaying various capacities of iron chaperone. Like just PCBP2 possesses the capacity of binding with the carrier systems like DMT1 as well as FPN1, in an iron-based method [60]. PCBP1 as well as PCBP2 both possess the capacity of transportation of iron to ferritin, with only PCBP1 being foundational in ferritinophagy, that constitutes a recycling event [61], where iron- ferritin complex gets engulfed by the nuclear receptor coactivator4(NCOA4) as well as is given direction towards the autophagosome [62].
Both PCBP1 as well as PCBP2 expression in pancreatic β cells have been demonstrated, however their particular part in iron tackling as well as if they are further implicated in transportation of iron to intracellular organelles as well as FeF proteins has not been proved in this cell kind.
2.4 Exchange of iron with organelles
Despite the observation of iron with practically each intracellular organelle, it is the mitochondria that is believed to be the major storage house with regards to iron cellular metabolism.
Actually, they represent the main station where iron storage in addition to utilization of iron occurs. Key generation of heme as well as iron-sulphur (Fe-S) clusters of electron transport proteins occurs amongst them.
The iron exchange with mitochondria is believed to be modulated via DMT1 as well as the traditional mitochondrial iron transporters mitoferrin (Mfrn)1 as well as 2 [63], as the one that is thought to be the 2nd greater particular with respect to non- erythroid cells [64]. Besides that, lipocalin (LCN) protein 2 is further implicated in this event of chaperone protein [6,65]. The continued expression of DMT1 has been the observation in case of HK293 that is the transporter existent at the outer cellular mitochondrial membrane (OMM) [65] in addition to implicated in, Fe 2+ as well as Mn2+ uptake [67]. Mfrn1 as well as 2 make sure that iron gets shifted across the inner mitochondrial membrane, where utilization of iron takes place for heme generation in addition to Fe-S clusters bio generation or gets isolated by the mitochondrial ferritin (MTFT).
The generation of Fe-S clusters needs frataxin i.e. an iron mitochondrial chaperone whose expression occurs in islets along with β cells, besides getting induced under , hyperglycemic situations[68]. Friedrich’s ataxia(FRDA) is a neurological disorder involving neurodegeneration that occurs secondary to frataxin deficiency in persons afflicted by this disease, in which non neurological sequelae like diabetes or glucose intolerance occurs as well with the incidence, of 8-32%[69].Thus these patients, have a manifestation of iron overload in addition to β cells apoptosis, which further corroborates the association with impairment of Iron homeostasis along with diabetes.
Iron gets exited from the mitochondrial matrix, the ATP-binding cassette (ABC) transporter gene ABCT7 is thought to cause Iron efflux as Fe-S clusters. This posit is dependent on the action of yeast orthologue Atm1 [70], which possesses the capacity of transportation of glutathione harmonized Fe-S clusters that connects the mitochondrial as well as cytosolic Fe-S cluster congregating systems [71]. More recently, Pearson et al. [72], corroborated this substance particular relation, emphasizes the part of Mg-ATP in this event of transportation [72]. Furthermore, an extra mode for Iron efflux from the mitochondrial matrix might be the exportation of heme by particular transporters [73].
Moreover, Iron can get supplied to the mitochondria via direct connection with the other organelles. In the generating erythroid cells for which effectiveness of iron supply to mitochondria for aid in heme generation, direct iron supply from endosomes to mitochondria for a mode described ( ‘’as kiss as well as run’’) mode [74], where shift of the cation gets modulated has been detailed via the voltage dependent anion channel (VDAC1) or DMT1[67]. In view of the applicable part that gets played by [67] iron in insulin liberation, akin modes of iron supply to the β cells can be believed to be existent, with the knowledge that akin event having been detailed in the epithelial cell [75].
Regions where mitochondria as well as lysosomes come in close proximity that is not associated with mitophagy or breakdown of mitochondrial vesicles by lysosomes have further been detailed by the aid of high resolution microscopy [76].Validating the functional importance of these intricate contacts with regards to iron transport in case of erythroid progenitors ,where administration of lysosomal- transferrin to mitochondria gets modulated by the TfR2 isoforms ,if there is TfR2deficient state reduction of mitochondrial size along with heme generation results [77]. Moreover, in cases of fibroblast from patients afflicted by neurodegeneration with brain iron accrual, aberrations of mitochondrial function, besides reduction in proteolytic action of lysosomes have been seen [78], that pointed that another mode of intracellular iron trafficking that is dependent on the crosstalk amongst mitochondria as well as a lysosome.
Mitochondria correlated ER (endoplasmic reticulum) membranes (MAMs) might further be implicated in the Iron homeostasis of the cell. If Cisd2 is deficient (CDGSH iron-sulphur domain2) an Fe-S protein that resides on MAMs results in mitochondrial impairment, besides impairment, of Ca2+ homeostasis causing an insulin sensitivity reduction in case of adipocytes [79]. Intriguingly in yeast deletion of protein complex ERMES (endoplasmic reticulum mitochondria encounter structure) that bridges 2 organelles, decides an Iron deficiency reaction in Iron depleted situations as well a, that results in escalation of iron in the cell [80]. Moreover, vacuolar protein sorting 13 (protein (VSP13p) dominants mutants rescue the ERMES mutants, that results in repression of Iron deficiency reaction. Till now Identification of no transporters with regards to iron transportation to endoplasmic reticulum (ER) have got proved. The 2 Fe-2S protein iron-sulphur domain2 (Miner 1) that resides in ER other cell types besides has importance in ER intactness [81] might be implicated in this function [82].
2.5 Regulatory proteins for Iron metabolism
Since both iron deficit in addition to iron overload might be deleterious, in β cells, iron genes regulation occurs post transcriptionally controlled by iron regulatory proteins (IRPs), that is dependent on how much iron is available [83]. They are RNA binding proteins, which via binding to IRE sequences that are existent on the mRNAs of iron tackling proteins, manipulate their translation. Specifically in situations of Iron deficiency, IRP binding occurs with TfR1, DMT1 besides ferittin mRNA along with facilitates their translation, hence resulting in escalation of iron absorption along with its storage [83]. Simultaneously IRP results in repression of FPN1 translation, hence reduction in cellular iron liberation [84]. IRP 1 as well as IRP 2 both get expressed in β cells, with IRP 2 knockdown mice generate type2 diabetes mellitus (T2DM) in view of aberrant iron metabolism [85].
3.1 Requirements of iron with regards to normal β cells function
Expression of greater amounts of iron in addition to storage proteins gets displayed by β cells besides demonstrate, escalation of iron metabolism in contrast to alpha as well as delta cells . The reason being iron acts as a cofactor of various enzymes besides being a necessary constituent of Fe-S cluster proteins that are implicated in important functions that vary from i)insulin liberation to ii)β cells proliferation, as well as differentiation (figure 2).
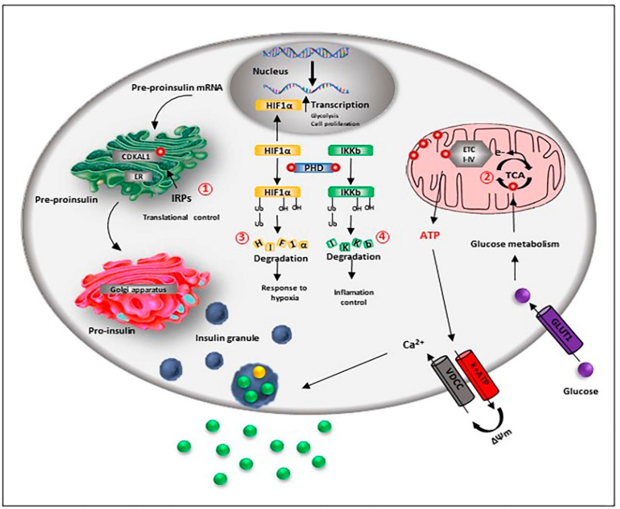
In agreement with this probability, mouse that have iron deletion reveal abnormal glucose stimulated insulin secretion, besides upregulation of transferrin surface expression in situations of glucose absence [86].
Insulin generation as well as liberation are significantly based on iron. Pro insulin translational allegiance in pancreatic β cells needs the action of the Fe-S cluster enzyme CDKAL1. This enzyme is implicated for the adenosine methylthiolation in the t RNA for lysine ,a modification that is the requirement for sustenance of the correctness of the codon recapitulation at the time of protein translation. On impairment of CDKAL1 function resulting in codon incorrect reading in addition to dysregulation of Proinsulin getting processed as well as liberation[85]. Intriguingly, mice with absence of IRP2 protein get the generation of T2DM secondary to subsequent iron deficiency, hence resulting in reduction of function of CDKAL1 [85].
Furthermore, iron requirement is there for the effectiveness of coupling of glucose metabolism along with insulin liberation. A crucial step for this mode is the glucose oxidation in the tricarboxylic acid (TCA) cycle for the generation of reducing equivalent, whose utilization by the respiratory chain results in the generation of the proton gradient that causes stimulation of ATP generation. The escalation of the ATP: ADP ratio as a result causes the closure of the ATP-sensitive-K+ channel (K ATP)- depolarization of the plasma membrane; the resulting opening of voltage-gated Ca2+ channels (CaV causing insulin liberation). Iron is implicated in the TCA cycle as the succinate dehydrogenase as well as aconitase that are implicated in the catalysis of the mandatory steps in the cycle are both iron based enzymes. ii)Furthermore, direct regulation of ATP generation is caused by iron, since Fe-S cluster protein is part of complex I, II, III as well as IV of the mitochondrial respiratory chain. In agreement with this probability in case of Ins-1 E β cell-line, silencing of ZIP 14 resulting in reduction of iron transportation into cells in addition to down regulation of numerous metal binding proteins, like the cytoplasmic iron detecting protein aconitase 1(ACO1) as well as ribosomal mitochondrial proteins, impacting the Oxidative phosphorylation along with liberation of insulin [45].
ii)Moreover indirect regulation of iron with regards to modulation of liberation of insulin occurs via the generation of ROS. Numerous steps in the insulin liberation possess sensitivity to the redox balance, like the depolarization of the plasma membrane; that gets initiated by the closure of the ATP-sensitive-K+ channel gets corroborated by the stimulation of the nonspecific cation channels (NSCC) [87]. Moreover, at the time of granule fusion the activity of voltage-gated Ca2+ channels (CaV) get magnified by the liberation of calcium from the ER, via a ROS based activation of the ryanodine receptor 2 [33,88].
iii)Furthermore, pancreatic β cells physiology gets controlled by iron, like proliferation, differentiation as well as survival. In the form of a cofactor of the HIF prolyl as well as asparaginyl hydroxylase (PHD), iron regulates the hypoxia inducible factor 1 alpha (HIF 1-α) breakdown besides participation in the β cells reaction to low oxygen situations [89]. During normoxia, PHD causes hydroxylation of HIF 1-α, hence resulted in its breakdown; whereas at the time of hypoxia or iron absence, PHD become inactive with HIF 1-α translocating to the nucleus as well as causing regulation of numerous genes transcription that are implicated in the control of the anaerobic glycolytic pathway. Consequently the modification of cellular metabolism occurs, with the cells moving from a proliferation phase towards a resting phase .In validation of this part absence of iron in view of lysosomal impairment, resulted in the stimulation of the HIF 1-α signaling with a resultant proliferation becoming dysfunctional [90].
iv)Via its association with HIF 1-α, iron might further impact the control of β cells working that gets modulated through the circadian clock mode. The metabolism of glucose in addition to insulin liberation are controlled by this mode in the β cells [91], along with interference with these circadian modes is implicated in the generation of T2DM in rodents along with humans [92,93]. Actually, a reciprocal crosstalk amongst the clock genes away the HIF 1-α transcriptional programming appears clear ,having the knowledge that HIF 1-α possesses the capacity of binding the promoter area of the clock genes in addition to regulation of their transcription, at the minimum chances in skeletal muscle cells [94], as well as on the other hand HIF 1-α by itself represents a direct target for transcription by the orthologue of Clock gene, NPAS2,in Hepatocellular carcinoma(HCC)[95]. Moreover, various iron associated genes as well as PHD get regulated at transcription level by the clock genes [96], which pointed that a circadian control of iron homeostasis is existent, which possesses the probability of being significant in the regulation of tissue particular metabolic reprogramming, that is dependent on oxygen as well as fuel existence. Intriguingly, alterations in Clock, NPAS2, as well as a Baml1 expression have been seen amongst newborns as well as adult rat islets, as acquiring circadian regulation of insulin liberation aids immature islets, possessing the properties of amino acid stimulated insulin bio generation as well as liberation, for attainment of the mature capacity of insulin liberation as a reaction to escalation of glucose amounts [97].
An appropriate consumption is further essential in β cells for regulation of inflammation in the form of iron based PHD hydroxylates, besides inactivation of the inhibitor of κB kinase (IKKb), that is an upstream significant controller of the nuclear factor κB(NFκB) pathway, that is the biggest proinflammatory pathway in β cells [98].
More recently, the results have pointed that probably there is implication of iron in β cell differentiation. Actually, TfR1, amounts, transferrin bound iron uptake as well as ferritin transcripts get upregulated in early post-natal weeks of the β cell maturation [86].This finding pointed that an escalated need of iron precisely at the time of metabolic shifting from aerobic glycolysis to Oxidative phosphorylation, that is essential for β cell maturation[99].As documented earlier ,various iron based proteins are needed for the maintenance of action of the totally mature β cells that are functional .
The biophysical properties of the extracellular milieu might impact the cell differentiation as well as survival of human islets of Langerhans along with β cells as demonstrated by Galli, et al [100].Their observation was that the extracellular matrix[ECM), nanotopography, through a mechanotransduction pathway ,that implicates a mechanosensitive integrins with reorganization of the actin cytoskeleton along with alterations in nuclear architectonic, that stimulates a particular transcriptional program that is essential for the metabolic adjustments of the cell towards the new surroundings. This reaction gets modulated by modifications of the mitochondrial action as well as dynamics besides implicated in the inter action of mitochondria with other organelles like lysosomes as well as ER, where iron gets exchanged [74,80] that further posits a probable part of iron with regards to this signaling.
3.2 β cells impairment occurs secondary to iron overload
A normal iron amount is needed for appropriate β cells function, however its escalation results in toxicity based on generation of ROS in addition to enhancement of stimulation of the Oxidative pathways (figure 3).
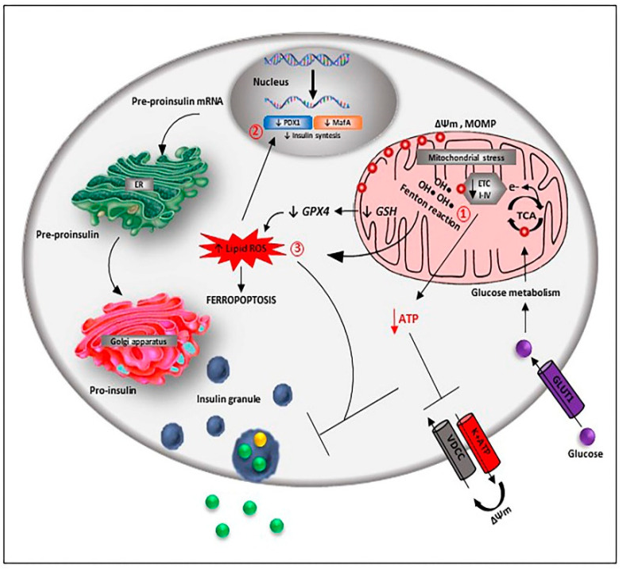
Hereditary haemochromatosis (HH) models have demonstrated, that iron accrual influences greater β cells in contrast to α cells or δ cells [101], possibly in view of the reduction of the ROS detoxification in this cell subkind.
Variety of modes have been posited mechanistically as a reason for iron toxicity; i) directly subsequent to intracellular iron overload, since in the form of a positively charged ion on gaining entry to mitochondria it can result in depolarization of the organelle membrane potential that has an impact on the electron transport chain (ETC), along with the energy liberation for insulin secretion [102]. Iron accrual can result in the development of ROS directly as well as indirectly. The active iron state (Fe 2+) results in oxidation of lipids in a Fenton’s like reaction with resultant ROS modulated DNA as well as protein oxidation, reduction in insulin generation as well as liberation, besides apoptosis as seen in the homeostatic iron regulator (Hfe) knockout mouse model of HH[103]. Noticeably other robust iron overload models that impact hepcidin expression like Hamps as well as Hjv knockout along with the hepcidin resistant model that possesses p.C326S mutation in ferroportin, despite manifesting pancreatic iron collection, do not display liver disease or endocrine abnormalities, possibly in view of higher Oxidative stress damage of mouse models[104]. The pancreatic as well as duodenal homeobox1(PDX1), in addition to V-Maf avian musculoaponeurotic fibrosarcoma oncogene homolog (Maf A), are 2 key transcription factors implicated in the regulation of insulin gene expression are the targets for ROS [105], along with reduction of hepcidin expression in the MIN6 cells with resultant hampering of insulin generation through iron overload along with reduction of PDX1 expression [106].
Furthermore, reduction of the β cells oxidant defenses takes place by iron overload via the hampering of the enzymes implicated in detoxification of ROS, like the, Mn2+ uptake as well as Mn2+ based SOD action [107].
Ii}An alternative mode of iron overload influence might be on the β cell function, besides survival through amylin. Misfolding along with accrual of Human islet amyloid polypeptide(HIAPP)- in the ECM as well as within β cells observation occurred studies in the postmortem pancreas of subjects afflicted by DM [108],where this polypeptide demonstrated cytotoxic action that was secondary to the disturbance of the cell membrane, disconcerted ion homeostasis ,ER stress , mitochondrial injury along with impairment in addition to Oxidative stress[rev in109].Interestingly iron has been demonstrated to result in escalation of amylin amyloidal beta sheet generation, that causes stimulation of the precipitation of amyloid aggregates [110]. Moreover, in the form of amylin, generation of a Heme Aβ and Heme- HIAPP complexes which might cause resultant generation of H2O2 through Oxidative stress [111,112], hence facilitation of β cells failure.
iii)More modes by which iron aids in the impairment of β cells failing along with demise is Ferroptosis which represents a nonapoptotic form of cell demise that occurs secondary to iron accrual. Initial observation of Ferroptosis was seen in cancer cells that had received treatment with glutamate/cystine exchanger (Xc) inhibitor elastin [113]. The properties of this are lipid ROS collection in view of hampering of glutathione peroxidase (GPx), by glutathione (GSH)deletion. Glucolipotoxic situations in mouse have been documented to cause escalation of β cell Iron import along with cytosolic ROS generation [114]. Moreover, pharmacologic hampering of GPx4 generation has been demonstrated to cause glucose modulated β cell impairment in vitro[115],where substances that cause induction of ferroptosis have been documented to interfere with the in vitro viability of human pancreatic islets along with their [116], function with the antidiabetic, quercetin has been demonstrated more recently to cause reduction in the ferroptotic injury in the pancreatic β cells of T2DM mouse models [117]. In the 1st instance Davalli et al. [118], could yield proof that escalation of extracellular amounts of glutamate might imply an insult for the β cells . Interestingly, the action of glutamate was not modulated by the stimulation of ionotropic receptors, however, by glutamate stimulated Oxidative stress that was correlated with the changes the glutamate/cystine exchanger (Xc) action, GSH) deletion in addition to enhancement of lipid peroxidation, a mode that is akin to ferroptosis [118].
More recently, fratraxin has further been attributed to be a controller of ferroptosis. In case of human fibrosarcoma HHT-1080 cells [119], repression of fratraxin expression causes acceleration of erastin stimulated cell demise, escalation of iron collection, lipid peroxidation along with mitochondrial injury processes that could get reversed by overexpression of fratraxin or pharmacological hampering of ferroptosis. As per proof with regards to this stimulation of a ferroptotic pathway of cell demise got derived from FRDA models ,like primary patient- obtained fibroblasts murine fibroblasts that possess FRDA correlated mutations along with fratraxin knockin/knockout murine fibroblasts[120].
Recently, it has been corroborated that failing of β cell in case of T2DM might be correlated with cell dedifferentiation even [121]. Epigenetic modifications or alterations in transcription factor action as well as /or associated difference in RNA or protein amounts might decide the depletion of β cells gene expression or the upregulation the genes that do not get expressed in the natural course in the mature β cells like the ones that get expressed in the islet’s progenitors or the other islets cell kinds [121]. Intriguingly, the Jumonji C-domain(JMJD6) possessing Histone arginine demethylase, that represents an epigenetic controlling enzyme which is iron based[122], besides in Hepatocellular carcinoma specimens a shifting of expression from TfR2 to TfR1 as well as overexpression have been correlated with tumor dedifferentiation, in addition to prognostically is poor [[141],that suggests the probability that dedifferentiation might be further associated with changes in the iron metabolism.
4. Implication of Impairment of Iron homeostasis in Diabetes
There has been corroboration that a direct association is existent amongst Impairment of Iron metabolism as well as diabetic situations. Earlier emphasizing on the escalation of ferritin in subjects with T2DM can be the observation [19], in addition to subjects with Metabolic Syndrome [123]. Escalation of the incidence of type2 diabetes mellitus (T2DM) (varying from 20-60%) has been visualized among patients with primary or secondary iron overload caused by Hereditary haemochromatosis (HH) or thallasemia in view of pancreatic β cells Impairment along with IR [124]. Furthermore, patients impacted by aceruloplasminemia, that represents an auosomal recessive condition in possession of the properties of absence of ceruloplasmin ferroxidase generation with accrual of iron in the brain along with liver, demonstrate diabetes mellitus generation as well[125], in addition to persons afflicted by Friedrich’s ataxia ,that is the neurodegenerative condition that occurs secondary to the deficit of the mitochondrial iron chaperone fratraxin [69].
Minimal iron overload also that is under the amounts that is classical of haemochromatosis or other iron storage abnormalities, has been correlated with the escalation of risk of generation of Gestational Diabetes mellitus [126,127], besides that of non-alcoholic fatty liver disease INAFLD) in the existence of Metabolic Syndrome [128].
Obesity, metabolic stress along with T2DM possess the properties of changed iron homeostasis; leptin deficiency in genetically obese (ob/ob) mice where generation of both Obesity along with T2DM occurs, demonstrate escalation of iron absorption along with getting retained [129]. Hyperglycemia has been illustrated to cause enhancement of DMT1 expression in microvillus membrane along with escalation of intestinal iron uptake in case of streptozocin (STZ), - induced diabetic mice along with brush border residing DMT1 have been the observation in intestinal biopsies in humans with T2DM [130]. Shu et al [106], documented a glucotoxicity- induced reduction in hepcidin expression, which resulted in failing of β cells by causing up regulation of TfR1 along with DMT1 that was followed by subsequent iron overload [106]. Frataxin which is the iron chaperone protein reduction has been the observation in islets from T2DM donors along with in the humanized model of frataxin deficiency FDRAYG8R, that results in iron overload in the mitochondria along with β cell Impairment [68].
In agreement with this an etiological part of iron in diabetes mellitus, it has been documented that dietary restriction causes enhancement of β cells function, along with glucose tolerance in (ob/ob) mice [125], along with phlebotomy has been demonstrated to result in enhancement of insulin sensitivity, insulin liberation along with glucose control in T2DM [132], despite clinical results that have been collected thus far are not conclusive with regards to this approach on iron letting or deletion [133].
β cells possess requirement of iron for their appropriate function. In view of that they have possession of numerous proteins that are implicated in the tackling of this ion, like the importers of iron, TfR1 along with DMT1 along with the iron storage proteins ferritin. The insight we do not possess, in the context of why alterations of proteins occur at the time of hyperglycemia, obesity, along with T2DM thus causing iron accrual that via ROS generation, results in Impairment of insulin generation, liberation along with apoptosis that aids in the generation of T2DM along with propagation. Intriguingly, certain drugs that have been posited for the therapy of diabetes mellitus demonstrate a definite action on iron homeostasis in the β cells .Like in the KIKO mouse model of FRDA, Exenatide that is an incretin mimetic has been recently corroborated to result in enhancement of glucose homeostasis by causing escalation of insulin liberation along with result in reduction of Oxidative stress via the induction of frataxin along with Fe-S cluster protein expression [134].Akin to that thiazolidenediones avoid mitochondrial iron collection [135] that further emphasizes on the association among iron homeostasis regulation in addition to conservation of β cells function.
Of the most significant factors that are implicated in the etiopathogenesis that influence pancreatic β cells function along with survival is Oxidative stress, with iron working as a catalyst in the generation of ROS by the Fenton’s reaction that might be a pointer to be a modulator of this event .The association among iron control impairment, in addition to failing of β cells has been proved with any aberrations in iron storage along with chaperone proteins has got correlated with diabetes generation or in case of diabetic situation .The query that requires clarification is if β cell sensitivity towards iron overload is secondary to low antioxidant ability of these cells or this in ability might result in enhancement of iron accrual in hyperglycemic situation along with /or hyperlipidemic situation. Getting further insight with this future exploration might aid in defining the iron regulation dependent anti diabetic strategies with the Objective of β cells preservation. Furthermore, in situations where anticipate this iron overload with any abnormalities of iron storage protein or chaperone proteins one can attempt avoidance of generation of T2DM like in FRDA having the knowledge of escalation of IAPP etc.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
