
Review Article | DOI: https://doi.org/10.31579/2690-1919/256
1 Department of Basic Medical Sciences, Inaya Medical College, Riyadh, Saudi Arabia.
2 Department of Physiology, Faculty of Medicine, Sabratha University, Libya.
3 Department of Biochemistry, Faculty of Medicine, Sabratha University, Libya.
*Corresponding Author: Azab Elsayed Azab, Department of Physiology, Faculty of Medicine, Sabratha, University, Libya. Email: azabelsaied@yahoo.com
Citation: Ata S.I. Elsayed, Azab E. Azab and Khawla A. Etwebi. (2022). An Overview of Oxytocin: Chemical Structure, Receptors, Physiological Functions, Measurement Techniques of oxytocin, and Metabolism. J. Clinical Research and Reports, 11(4); DOI:10.31579/2690-1919/256
Copyright: © 2022, Azab Elsayed Azab. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 06 June 2022 | Accepted: 20 June 2022 | Published: 30 June 2022
Keywords: oxytocin; structure; receptors; physiological functions; measurement techniques; measuring receptors; metabolism
Background: Oxytocin is a peptide hormone comprising 9 amino acids. It is produced in the hypothalamus and stored and secreted by the posterior lobe of the pituitary gland and synthesized in other organs such as the uterus, ovaries, placenta, heart, blood vessels, skin, kidneys, and testis. Receptors of oxytocin are present on myoepithelial cells, heart, blood vessels, macrophages, thymus, pancreas, kidneys, and adipocytes.
Objectives: The current review aimed to give highlight the oxytocin structure, receptors, physiological functions, measurement techniques, and metabolism. Oxytocin is a small peptide that consists of nine amino acids in a six–amino acid ring formed by cysteine bonds and a three–amino acid tail with a terminal amine. It is synthesized in brain regions that are critical to behavioral and physiologic homeostasis. Oxytocin is involved in uterine contraction during labor and ejection of milk during breastfeeding and plays a role in social behavior, emotions love and affection, the period after childbirth, and metabolic functions. The action of oxytocin in facilitating human bonding and social relation is well known. The effects of Oxytocin on metabolism and food intake suggesting its potential effects in treating obesity. The half-life time of oxytocin in the brain is triple as long as its half-life time in the periphery. The sensitivity and density of oxytocin receptors increase during labor. After birth, the neonatal baby sucks on his mother's breast, causing the release of milk by stimulating hypothalamic neurons to produce oxytocin. Oxytocin neurons have been heavily implicated in mediating sexual behavior in both humans and animals. The social memory was enhanced by central oxytocin administration in male rats. The action of oxytocin affects social memory in multiple brain regions, including the ventral hippocampus, amygdala, olfactory bulb, and lateral septum. Oxytocin neurons may mediate MC4R-driven sexual behavior in male mice. MC4R signaling in oxytocin neurons permits ejaculation. A decreased latency to ejaculate in rabbits and rats after administration of oxytocin. The effect of oxytocin receptor ligands on the ejaculatory response may be due to the modulation of dopamine serotonin neurotransmission. Oxytocin lowers the threshold for the initiation of maternal behavior but is not involved in its maintenance. The oxytocin and the melanin-concentrating hormone (MCH) systems may interact to modulate maternal behavior. Oxytocin regulates maternal- or mating-regulated mood. Initial measurements of oxytocin by using radioimmunoassay and bioassays suggested that oxytocin concentration in blood is very low, 5 pg/ml, with small increases as pulses,15 pg/ml, during lactation and uterine contractions. Variations in oxytocin concentration, especially in rapid response to specific experiences, such as anticipation of breastfeeding, sexual stimulation, exercise, affiliative social contact, and psychologic stress. Oxytocin is rapidly removed from the plasma by the liver and kidney. Oxytocinase activity increases throughout pregnancy and peaks in the plasma, placenta, and uterus near term. It is also expressed in mammary glands, the heart, the kidney, and the small intestine. Lower levels of activity can be found in the brain, spleen, liver, skeletal muscle, testes, and colon. The plasma half-life of oxytocin ranges from 1 to 6 minutes. The half-life is decreased in late pregnancy and during lactation.
Conclusion: It can be concluded that oxytocin is a peptide hormone that is synthesized in brain regions and other organs and posse's receptors in many organs. It plays a role in social behavior, emotions love, and affection, the period after childbirth, and metabolic functions. Its potential effects in treating obesity. The half-life time of oxytocin in the brain is triple as long as its half-life time in the periphery. Oxytocin neurons may mediate MC4R-driven sexual behavior. Variations in oxytocin concentration, especially in rapid response to specific experiences. Oxytocin is rapidly removed from the plasma by the liver and kidney. The plasma half-life of oxytocin ranges from 1-6 minutes. The half-life is decreased in late pregnancy and during lactation.
All animal cells are exposed to a variety of extracellular signals that need interpretation and translation into the appropriate response to the surrounding environment. These signals can be soluble chemical substances factors generated locally or from distant glands, such as hormones. Achieving this ligand cells maintained a diversity of cell surface receptors responding specifically to this ligand. These receptors are classified into many families according to the way in which they generate the intracellular signals pathway to give the particular responses and functions. The activity of any receptor can be modulated by other signaling pathways in different ways, generating the flexibility needed for such a complex system. the major classes of receptors included G protein-coupled receptors, ligand-gated ion channel receptors, tyrosine kinases receptors, integrins, and cytokine receptors [1].
Oxytocin hormone is produced in the hypothalamus and stored and secreted by the posterior lobe of the pituitary gland. oxytocin is involved in uterine contraction during labor and ejection of milk during breastfeeding, it also has many effects on social behavior, emotions love, affection, and metabolic functions. Many studies were conducted on its effects in weight loss by decreasing food consumption and increasing energy consumption. The effects of Oxytocin on metabolism and food intake suggesting its potential effects in treating obesity [2].
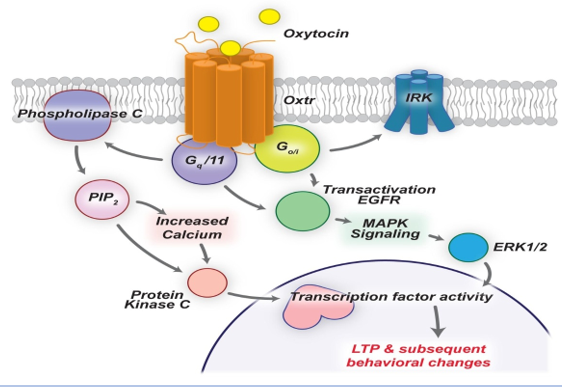
Oxytocin is a peptide hormone comprising 9 amino acids produced mainly in the brain and many peripheral organs. It binds to its receptor, which is G protein-coupled receptor. Synthesis of oxytocin occurs in paraventricular and supraoptic nuclei in the hypothalamus. The magnocellular neurons extend axons to the posterior lobe of the pituitary gland. The hormone is secreted into the circulation and cannot back to the brain due to the presence of the blood-brain barrier. Other axonal connections exist between the paraventricular oxytocin-producing neurons and regions of the brain, brainstem, and spinal cord. Parvocellular neurons in the paraventricular nucleus connect to the nuclear bed of the ventral tegmental area, stria terminalis, and the nucleus accumbens which are involved in eating behavior and reward behavior regulation. G protein-coupled oxytocin receptors are expressed throughout the brain in structures such as the hypothalamus, amygdala, olfactory nucleus, and anterior cingulate cortex in the limbic system. The interaction between oxytocin and other neurotransmitters, such as serotonin, increases oxytocin concentrations. The reaction of dopamine and oxytocin regulates the activity of the reward circuitry in the brain (Figure 1).
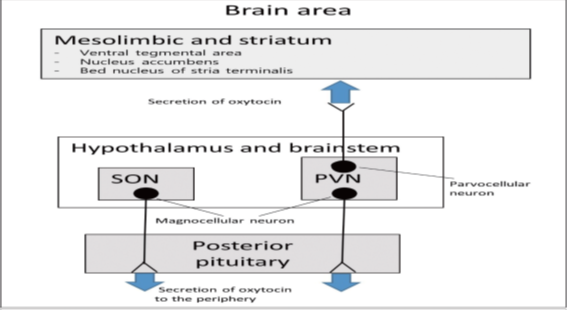
SON: supraoptic nucleus; PVN: paraventricular nucleus [3].
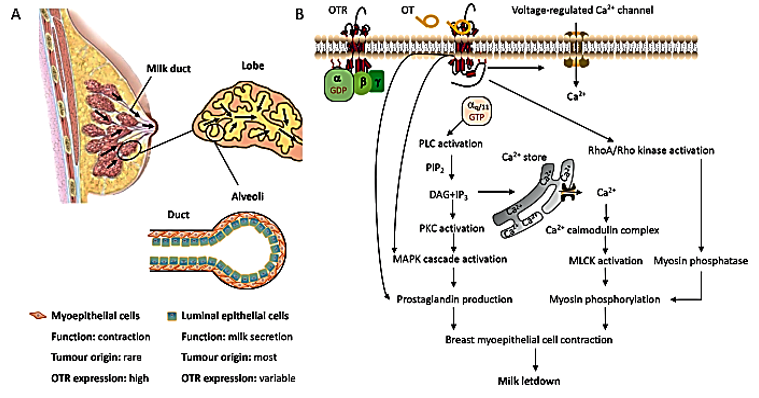
Also, oxytocin is synthesized in other organs such as the uterus, ovaries, placenta, heart, blood vessels, skin, kidneys, and testis. Receptors of oxytocin are present on myoepithelial cells, heart, blood vessels, macrophages, thymus, pancreas, kidneys, and adipocytes.The receptors of oxytocin are involved in uterine contractions, nitric oxide production, and lipolysis through G protein coupling and C-beta pathway activation. Secretions of oxytocin in GIT are associated with paracrine and autocrine effects.Positive feedback mechanisms were noticed from the peripheral or central administration of oxytocin, stimulating autoreceptors in the magnocellular neurons in the supraoptic nucleus.Oxytocin receptors are found throughout the central nervous system and peripheral regions, providing a clue for the regulation of metabolism and food consumption. In addition to the anterior pituitary gland, oxytocin receptors have been found in pancreas, gastrointestinal tract, adipocytes, and studies have reported their association with eating behaviors (Figure 2) [3].
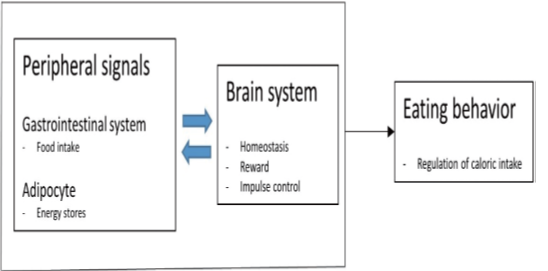
The current review aimed to give highlight the oxytocin structure, receptors, physiological functions, measurement techniques, and metabolism.
Chemical structure of oxytocin
Oxytocin is a small peptide that consists of nine amino acids in a six–amino acid ring formed by cysteine bonds and a three–amino acid tail with a terminal amine. Oxytocin is synthesized in conjunction with carrier proteins (neurophysin 1). The gene for oxytocin is located on human chromosome 20, adjacent to the gene for vasopressin. These genes lie on opposite strands and have opposite transcriptional orientations [4].
It is synthesized in brain regions that are critical to behavioral and physiologic homeostasis. Different cells in specific brain regions produce this hormone, including the supraoptic nucleus and paraventricular nucleus in the hypothalamus. The neurons of the paraventricular nucleus termed magnocellular neurons, synthesize oxytocin and extend their processes to the posterior lobe of the pituitary gland to release it into circulation. The hypothalamus paraventricular nucleus is a major site of integration and convergence for neural communication relating to stress with effects on the hypothalamus-pituitary-Adrenal axis and autonomic nervous system functions. Oxytocin is co-localized in at least some of its cells of origin with corticotropin-releasing hormone, which in turn regulates the hypothalamus-pituitary-Adrenal axis. Corticotrophin-releasing hormone actions also have been implicated in some of the detrimental effects of chronic stress. Oxytocin may be co-released with Corticotrophin releasing hormone or vasopressin as an adaptive response to a variety of challenges and facilitating coping (Figure 3) [5].
Conventional arrow = agonist; circle-tipped arrow = positive allosteric modulator; block-tipped arrow = antagonist [6].
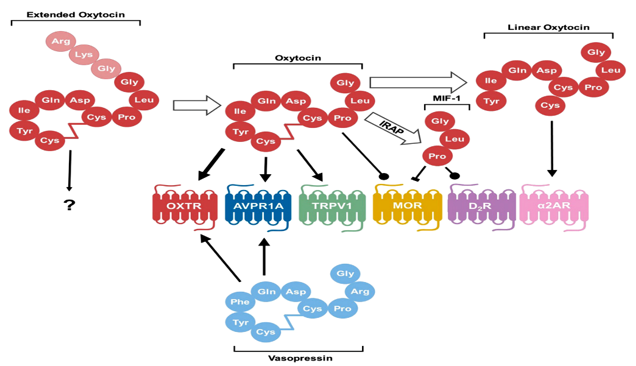
After conversion from the prohormone form, oxytocin exists in an extended form with three extra amino acids. The conversion from extended oxytocin to oxytocin consisting of nine amino acids occurs with the maturation of the hypothalamus in neurotypical individuals. It is not known how this extended form interacts with receptors. In their canonical nine–amino acid forms, both oxytocin and vasopressin bind and act as agonists to both oxytocin receptors and arginine vasopressin receptor1A, although oxytocin has a higher affinity for oxytocin receptors than the arginine vasopressin receptor1A, as denoted by the thicker arrow. Oxytocin also acts as an agonist to the pain-sensing transient receptor potential vanilloid-1 (TRPV1) receptor and as a positive allosteric modulator at the MOR. After degradation by IRAP, the C-terminal tail is cleaved from oxytocin to form MIF-1, which can both inhibit MOR and act as an allosteric modulator on the D2 subtype of dopamine receptors (D2R). Oxytocin can also be degraded by other as of yet unspecified peptidase activity into a linear form that stimulates activity of the a-2 type adrenoreceptors (a2ARs) [6].
Oxytocin receptors
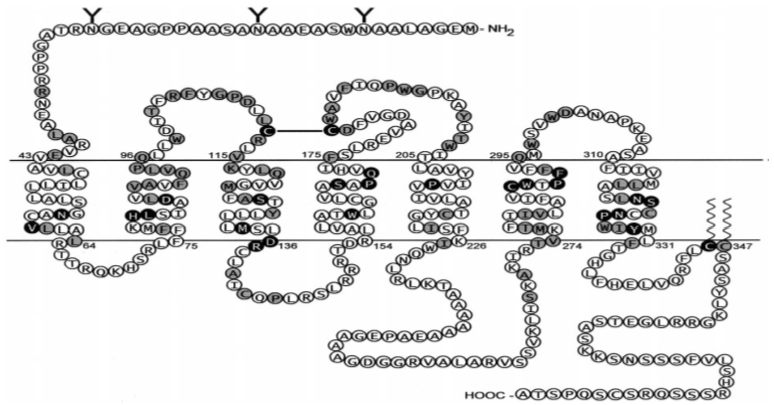
The receptor of oxytocin is a member of the G protein-coupled receptors family, rhodopsin-type (class I). The seven transmembrane α-helices are most highly conserved among the G protein-coupled receptors family members. Conserved residues among the G protein-coupled receptors may be involved in a common mechanism for activation and signal transduction to the G protein. The switching from the inactive to the active conformation is associated with a change in the relative orientation of transmembrane domains 3 and 6, which then unmasks G protein binding sites. In the class, I G protein-coupled receptors, and Asp in transmembrane domain 2 (Asp-85 in human oxytocin receptor and a tripeptide v (E/D RY) at the interface of transmembrane 2 and the first intracellular loop are believed to be important for receptor activation. With respect to Asp-85, this was confirmed for the human oxytocin receptor. When Asp-85 is exchanged by the residues Asn, Gln, or Ala, agonist binding and signal transduction of the receptor becomes impaired. Mutations at the conserved tripeptide motif DRY (DRC in the case of the oxytocin receptor) result in an either inactive or a constitutively active oxytocin receptor. The cysteine residues in the first and second extracellular loops are highly conserved within the G protein-coupled receptors and are probably connected by a disulfide bridge. Two other well-conserved Cys residues reside within the COOH-terminal domain. Most likely, they are palmitoylated as demonstrated for the vasopressin (V2) receptor and other G protein-coupled receptors, and anchor the cytoplasmic tail in the lipid bilayer. However, for the vasopressin (V2) receptor as well as for the rat oxytocin receptor, elimination of palmitoylation sites by mutagenesis failed to produce significant alterations in receptor function. The oxytocin receptor has two (mouse, rat) or three (human, pig, sheep, rhesus monkey, bovine) potential N-glycosylation sites (N-X-S/T consensus motif) in its extracellular NH2-terminal domain. For the “core” oxytocin receptor, a molecular mass of; 40–45 kDa can be calculated on the basis of the amino acid sequence derived from the known cDNA sequences of several species. In photoaffinity labeling experiments using myometrial membranes obtained from guinea pigs during late pregnancy, a 68- to 80-kDa protein was specifically labeled by a photoreactive oxytocin antagonist. Deglycosylation of the photo labeled receptor with endoglycosidase F gave rise to a protein with 38–40 kDa. Similarly, in the same tissue, a 78-kDa protein was labeled by a photoreactive vasopressin analog. In contrast, in membranes from rat mammary gland and rabbit amnion cells, photoreactive oxytocin analogs are specifically incorporated into a 65-kDa binding protein. It is possible that the different molecular masses for the myometrial versus the mammary gland and amnion oxytocin receptor are due to differential glycosylation patterns. With the assumption of a mass of; 10 kDa for a typical glycosylation core, all of the potential glycosylation sites could be occupied by glycosylation moieties. Recombinant deglycosylation mutants of the human OT receptor have been created by site-directed mutagenesis by exchanging Asp for Asn in positions 8, 15, and 26. The deglycosylated receptors were highly expressed in HeLa cells and showed unaltered receptor binding characteristics. Thus, receptor glycosylation appears not to be necessary for proper expression and has no effect on the functional properties of the receptor [7].
One oxytocin receptor is only described, the gene for it is located on chromosome 3p24–26 in humans [8]. The oxytocin receptor gene encodes a G-protein–coupled receptor with a seven-transmembrane domain. The same oxytocin receptor is present in nervous tissue and other parts of the body as the uterus, GIT, and breast. oxytocin affects numerous different G protein-coupled receptors3). Oxytocin can bind to receptors that were identified in vitro, such as oxytocin receptors, but also to receptors of vasopressin (AVPR1A, AVPR1B, AVPR2) [9]. There are several other target receptors for oxytocin’s action as its action as an agonist on the pain-sensing transient receptor potential vanilloid-1receptor, where it attenuates pain sensation. Oxytocin also acts as a positive allosteric modulator of the m-opioid receptor (Figures 4 & 5) [10].
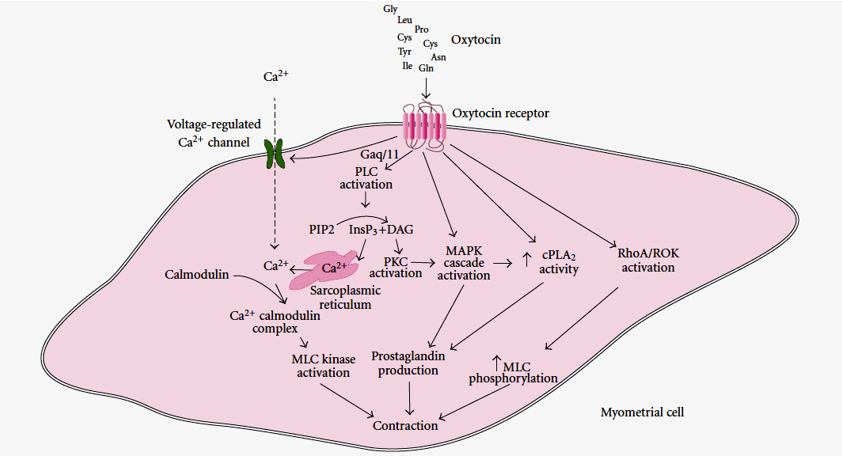
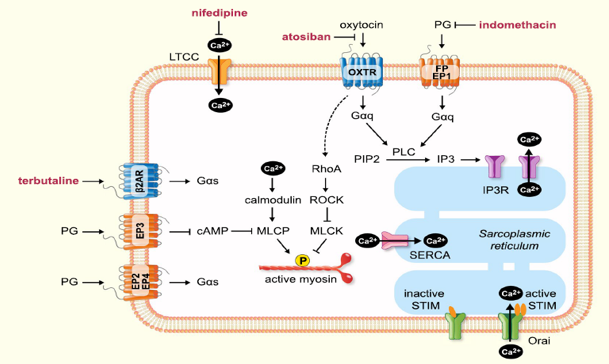
The binding of oxytocin to its receptors activates Gαq/11 and then phospholipase C, which hydrolyses phosphatidylinositol 4,5-bisphosphate (PIP2) to InsP3 and diacylglycerol. inositol 1,4,5- triphosphate (InsP3) causes the release of Ca2+ from the sarcoplasmic reticulum, while DAG activates PKC. Gαq/11 also causes activation of voltage-regulated Ca2+ channels and Ca2+ entry into the cells. Ca2+ binds to calmodulin, and the Ca2+-calmodulin complex activates myosin light-chain (MLC) kinase, resulting in myometrial contraction. Both oxytocin receptors and protein kinases type C (PKC) activate the mitogen-activated protein kinase (MAPK) cascade, while oxytocin receptors and MAPK result in increased cPLA2 activity. Increased cPLA2 activity and MAPK activation result in prostaglandin production, which also contributes to the contractile effect. The activation of the RhoA associated protein kinase (Figures 6 & 7) [11].
Physiological functions of oxytocin
Roughly 40% of paraventricular neurons which produce oxytocin project to central nervous structures such as the brain stem and pituitary gland, and 10% of these neurons connected to the brain stem have a role in the regulation of eating behavior via area postrema and nucleus tractus solitarius, the dorsal motor nucleus of the vagus nerve. The half-life time of oxytocin in the brain is triple as long as its half-life time the in the periphery, it is about 19 minutes in the brain and 6 minutes in the periphery, which means that, this neuropeptide acts as a relevant modulator of CNS function. Serotonin has been found to increase oxytocin concentrations while dopamine interactions with oxytocin can modulate the activity of the reward circuitry of the brain, this means that there is a relation between oxytocin and behavioral disorders such as depression, autism, and, eating disorders. Secretion of oxytocin in peripheral regions such as GIT submucosal and myenteric nerve plexus was also noticed, which augments its role in feeding and satiety behavior [14]. The effects of oxytocin on the uterus are well known. The sensitivity and density of oxytocin receptors increase during labor. After birth, the neonatal baby sucking of his mother's breast, causing the release of milk by stimulating hypothalamic neurons to produce oxytocin, causing contraction of myoepithelial cells of alveoli. Eating disorders are more dominant in females which speculates that because of the particular functional relevance of oxytocin in the female organism, changes in oxytocin pathways contribute to dysfunctional eating behavior patterns and that interventions aimed at these pathways hold a certain therapeutic action for this neuropeptide [14].
Social behavior
Animal studies on the effect of oxytocin on social behavior in a variety of mammals, including mice, rats, and sheep. These studies demonstrated oxytocin’s ability to induce the prosocial behavior of bonding, both pair-bonding and maternal bonding of mothers with their offspring. The pair-bonding behavior was measured by time spent in proximity of a partner versus a stranger, in which as the time spent with a partner was longer, means pair-bonding formation. Pair bonding can be induced in females. Injection of oxytocin in the intracerebroventricular region in the brain of
voles leads to the formation of pair-bonding among these animals, it can also be prevented by oxytocin receptor antagonists in the nucleus accumbens and prefrontal cortex of the brain. Maternal and offspring bonding has also been demonstrated also in voles. Individuals with social anxiety problems such as autism can struggle with forming and maintaining interpersonal relationships. The action of oxytocin in facilitating human bonding and social relation is well known, and future studies were needed for the possibility of treatment of social anxiety and autism by using oxytocin agonist drugs. Another aspect of social behavior the ability to recognize the individuals and differentiate between them, which is known as the social memory, was enhanced by central oxytocin administration in male rats, whereas using antagonists for oxytocin receptor blocks social memory in both male and female rats. The action of oxytocin affects social memory in multiple brain regions, including the ventral hippocampus, amygdala, olfactory bulb, and lateral septum. Studies on monkeys suggested the potential effects of oxytocin in promoting prosocial behaviors and enhancing social cognition in humans in spite of the complexity of social behavior in humans [15].
The role of melanocortin system and oxytocin neuropeptide in male sexual behavior
The dysfunction in sexual ability has effects on the quality of life in men. At least about 20% of men experience some form of dysfunction in their sexual forces during different periods of their life. The relation between age and decline in sexual function is well documented, this correlated to weight gaining and consequent cardiovascular diseases and diabetes with their complications. Although treatments with phosphodiesterase inhibitors drugs as sildenafil or other drugs for improving erection and sexual functions, responsiveness to these drugs are varying, and medications for other forms of sexual dysfunction are limited [16].
The targeting potential impairments of medications in brain circuits that underlie sexual function have newly recently garnered interest. For example, selective inhibitors for serotonin reuptake, which leads to an increase in serotonin in extracellular fluids, have been used in the treatment of rapid ejaculation. The agonists of dopamine as apomorphine were explored as sexual motivation and erectile dysfunction treatment. The melanocortin system has a role in erection function, and many drugs which targeted this system has been tested for erectile dysfunction treatment. Understanding the neurocircuitry mechanisms of sexual function may lead to more effective treatments [17].
Many clinical and preclinical studies support the involvement of the melanocortin system in sexual abilities and sexual actions. Melanocortin hormones, such as adrenocorticotropic hormone (ACTH), α-melanocyte-stimulating hormone (MSH), γ-MSH, and β-MSH, are produced from the proopiomelanocortin (POMC) polypeptide. These products bind to five different melanocortin receptors (MC1R, MC2R, MC3R, MC4R, MC5R) with different degrees of affinities. Since the MC3R and MC4R are the primary subtypes expressed by the brain, these two may mediate the central effects of melanocortin. MC4 receptors are G protein-coupled receptors that can couple with Gi/o, Gs, and Gq receptors and trigger a series of downstream pathways. The studies suggested that melanocortin receptors, MC4R, which have a role in satiety mediating, may mediate the sexual abilities and sexual actions. The administration of α-melanocyte-stimulating hormone, which is an agonist for MC3R and MC4R, has been found to lead to erection in mice and rats. At three to six months of age, MC4R null mice show decreased motivation to engage in sexual activity and take a longer time to reach ejaculation [18].
Male mice with insensitive POMC-producing neurons to insulin and leptin show reduced production in αMSH and MC4Rs expression. These mice showed a decrease in sexual motivation which was indicated by a reduction in mounting behavior during the initial sexual encounter; this phenotype is evident at four to six months of age, concurrent with increases in their weights and a decrease in insulin sensitivity [19].
POMC neurons project to nuclei that express Sim1, such as the supraoptic nucleus, paraventricular nucleus, and regions of the amygdala including the medial amygdala and basolateral amygdala. The study of Semple and Hill, [20] showed that genetically re-expressing MC4Rs only in Sim1 neurons of MC4RKO mice restores both intromission and ejaculation efficiency at age 6 months. The receptors MC4Rs in Sim1 neurons play a role in regulating feeding behavior; the weights of these mice were duplicated compared to control at the testing time. Therefore, it remains to be determined whether MC4Rs on Sim1 neurons directly regulate sexual function, or whether obesity is a necessary dysfunction factor [20].
Oxytocin neurons comprise a large portion of the Sim1 neuron population and are found in the supraoptic and paraventricular nuclei, downstream of POMC neurons. Oxytocin neurons have been heavily implicated in mediating sexual behavior in both humans and animals. These neurons are postulated to mediate the effects of MC4 receptor agonists on female sexual functions and partner preference. Semple et al., [17] hypothesized that oxytocin neurons may mediate MC4R-driven sexual behavior in male mice. They examined the ability of MC4Rs on oxytocin and Sim1 neurons in maintaining sexual function using mice, for 2-month-old, without obesity-related to age. their findings underline the feasibility of identifying specific neuronal targets for male sexual dysfunction treatment [17].
When the melanocortin 4 receptor (MC4R) is knocked out globally, male mice displayed low sexual desire, obesity, and difficulties in copulation; however, it is unclear whether these phenotypes are interdependent. To elucidate the neuronal circuitry involved in sexual dysfunction in MC4R knockouts, Semple et al., [17] in their study, re-expressed the MC4R in these mice exclusively on a subset of Sim1 neurons, namely oxytocin neurons (tbMC4Roxt mice) or Sim1 neurons (tbMC4RSim1 mice). The groups were compared to age-matched mice to observe the effects of obesity. In this experiment, young MC4R null mice had no deficits in erectile function or sexual motivation. However, MC4R null mice were found to have an increased latency to reach ejaculation compared to control mice, which was restored in both tbMC4RSim1 and tbMC4Roxt mice. These results indicate that the signaling of melanocortin via the MC4R on oxytocin neurons is important for normal ejaculation independent of the male’s metabolic health [17].
Oxytocin’s role in regulating erection and ejaculation has been studied in mice and rats. Oxytocinergic projections from the paraventricular nucleus to the hippocampus, medulla oblongata, and spinal cord facilitate penile erection. The results of Clement and Giuliano study, [21] revealed that MC4R signaling in oxytocin neurons permits ejaculation but argue against this pathway directly mediating erectile function. Systemic and intracerebroventricular (ICV) administration of oxytocin has been found to facilitate ejaculatory function, while intravenous (IV), ICV, and intrathecal administration of an antagonist of oxytocin receptor (GSK557296) have been cause inhibition in ejaculation. Similar studies have found a decreased latency to ejaculate in rabbits and rats after administration of oxytocin. The effect of oxytocin receptor ligands on the ejaculatory response may be due to the modulation of dopamine serotonin neurotransmission [21].
Genetic deletions of the peptide (MCH-KO) or its receptors (MCHR1-KO) result in mothering deficits similar to those seen in oxytocin receptor KO (OXTR-KO) mice. Both the melanin-concentrating hormone (MCH) and oxytocin lower the threshold for the initiation of maternal behavior but are not involved in its maintenance. Therefore, the oxytocin and MCH systems may interact to modulate maternal behavior. While only 4% of oxytocin neurons express MCHR1, OXTR mRNA is expressed in the majority of MCH neurons and only rarely in other neurons in the lateral hypothalamus (LH). Electrophysiological studies report that oxytocin interacts and selectively excites MCH neurons but not any other LH neurons [22].
The very selective action of oxytocin and the specific location of its receptors on MCH neurons suggests that these neurons may mediate or modulate some of the oxytocin actions on maternal behavior and emotion. The study by Sanathara et al. [23] showed that the MCH and oxytocin systems interact directly, nonetheless, whether oxytocin regulates maternal- or mating-regulated mood through oxytocin-MCH signaling is unknown. It is plausible to speculate that oxytocin may exert parts of its facilitating actions on mating and maternal responses through interacting with MCH neurons.
Measurement techniques of oxytocin
Oxytocin levels in bronchoalveolar lavage samples from asthmatic and healthy subjects were assayed in the study of Amrani et al., [24] using a competitive enzyme immunoassay, the method proceeds as follows: 100 μl of all standards and bronchoalveolar lavage samples were loaded in triplicate with 50 μl of blue conjugate antibody into each well, except the total activity and blank wells. The plate was sealed and incubated at 4°C for 24 hr. After incubation, wells were washed 3 times, and pNpp substrate solution was added and incubated at room temperature for 1 hr. Subsequently, the plates were read immediately at an optical density of 405 nm. The concentration of oxytocin in the samples was calculated based on the standard curve obtained with known concentrations of oxytocin.
Initial measurements of oxytocin by using radioimmunoassay and bioassays suggested that oxytocin concentration in blood is very low, 5 pg/ml, with small increases as pulses, 15 pg/ml, during lactation and uterine contractions [25]. Studies using immunoassay or mass spectrometry preceded by an extraction procedure continued to detect concentrations in the 5–30 pg/ml range. However, other methods, including mass spectrometry, have indicated that endogenous oxytocin may occur in blood at much higher concentrations greater than 500–1000 pg/ml. Studies on concentrations of oxytocin in tissues may give results of a particular understanding of the beneficial effects of this neuropeptide. For example, within the microenvironment of ovarian tumors, oxytocin levels were measured at 200 times higher than those in plasma. These high levels of oxytocin may reflect a response to the tumor with apparent benefits to reducing inflammation. But they also could be related to the absence within the tumor compared with the blood of peptide binding molecules, which, in turn, might affect the concentrations of oxytocin detected by antibody-based methods [26, 27].
However, many other works indicated that oxytocin readily engages in complexes at its disulfide bridge and that the vast majority of oxytocin in bio-samples evades detection using conventional approaches to measurement [28]. Importantly, these findings have been confirmed using mass spectrometry which is highly specific compared with immunoassay suggesting that earlier critiques of the high oxytocin concentrations detected by immunoassay may have been misguided. However, despite its advantages regarding specificity, mass spectrometry methods for measuring oxytocin are still in active development, and many researchers have faced substantial challenges in the reliable detection of oxytocin using this approach. Hence, it is important to keep in mind that although mass spectrometry is a very sophisticated approach, the verdict on the optimal measurement method is still being debated. Additional areas in which mass spectrometry measurement of oxytocin can be improved include the removal of interferents (e.g., phospholipids, which can suppress signals, resulting in vast underestimation of oxytocin concentrations), automation (including sample preparation) for higher throughput, and further development and maturation of high-sensitivity microfluidic separation systems [6].
Salivary measurements, possibly with fewer sources of variation, offer an alternative for the assessment of changes in oxytocin. Several studies have revealed reliable variations in oxytocin concentration, especially in rapid response to specific experiences, such as anticipation of breastfeeding, sexual stimulation, exercise, affiliative social contact, and psychologic stress [29, 30]. However, the relationships between concentrations of peptides in saliva, blood, and peptide concentrations in the brain nuclei are not yet well-understood [31]. The study of Yamamoto and Higashida [27] suggested that oxytocin is dynamically changing in biologic matrices, like blood. In addition, detecting the presence of binding molecules will be critical to accurate representations of the availability of these elusive molecules [27].
Measuring the Oxytocin Receptor
The measurement of oxytocin receptors is problematic because the pharmacological tools that are available for stimulating, identifying, or blocking receptors for oxytocin may not be sufficiently selective to allow easy manipulations or identification of these receptors in vivo. Cell culture or do not reflect the functional availability of these peptides, especially since the binding that occurs in vivo, including in tissues or blood, could be influenced by competing molecules that differ from one test to another, as in the case of microglial oxytocin receptors has been difficult to detect in vivo but appears to be significantly upregulated after immune challenges and stressors, especially when macrophages and microglia are challenged in vitro [27].
In the study Amrani et al. [24], Real-time PCR was performed to assess oxytocin receptor expression. In this study total, RNA was isolated, 1μg of total RNA was reverse transcribed at 37°C for 120 min followed by 25°C for 10 min. Forty ng of cDNA per reaction was used in the real-time PCR in the presence of Ampli Taq Gold DNA polymerase, the reaction was incubated for 2 min at 50°C followed by 10 min at 95°C. Then the reaction was run for 40 cycles at 15 sec, 95°C, and 1 min, 60°C per cycle. Assays-on-Demand™ primers and probes specific for oxytocin receptors were used in the PCR. The endogenous 18 S rRNA was measured and used to normalize all samples using the ΔΔCT method. Gene expression level of oxytocin receptor is expressed relative to 18 S and untreated samples in each stimulation study, respectively. At least 3 (often 6) replicates were run for each condition.
Drug discovery
Oxytocin plays a key role in adaptive processes associated with stress responses, tolerance reward, and memory. Through interactions with stress and brain reward systems, it is known to play a role in several psychiatric diseases, particularly those that involve altered social integration, such as addiction to drugs and alcohol. There is growing interest in the neuropeptide oxytocin as a potential therapeutic agent in the treatment of alcohol and drug abuse disorders. Accumulating preclinical evidence suggests that oxytocin administration affects the development of sensitization, tolerance, and withdrawal symptoms, and modulates numerous alcohol and drug-seeking and also alcohol and drug-taking behaviors. Further, there is some evidence to suggest that oxytocin may help to reverse neuroadaptations that occur as a result of the chronic taking of drugs and alcohol [32].
Early research in the drug discovery field indicated that administration of exogenous oxytocin can prevent the development of tolerance to opiates and ethanol, the induction of stereotyped, hyperactive behavior by drugs or ethanol, and the withdrawal symptoms associated with sudden abstinence from drugs and alcohol. Additionally, stimulation of endogenous oxytocin systems is underlying the prosocial and empathogenic effects of party drugs such as GHB (Fantasy) and MDMA (Ecstasy). Brain oxytocin systems exhibit profound neuroplasticity and undergo major neuroadaptations which resulted from drug use. Many drugs, including cocaine, cannabis, MDMA, GHB, opiates, and alcohol, cause long-term changes in markers of oxytocin function and this may be related to enduring deficits in social behavior in laboratory animals exposed to these drugs. Preclinical studies have illustrated a remarkable ability of exogenous oxytocin to inhibit stimulant and alcohol self-administration, alter associated drug-induced changes in glutamate, dopamine, and Fos expression in basal ganglia and cortical sites, and preventing of stress and priming-induced relapse to drug-seeking. Oxytocin, therefore, has fascinating potential to reverse the corrosive effects of long-term drugs abuse on social behavior and to inoculate against future vulnerability to addictive disorders. The clinical studies examining the effects of intranasal oxytocin in humans with drug abuse are eagerly awaited [33].
Oxytocin neuropeptide has a strong therapeutic potential for the treatment of a number of psychiatric disorders as social impairments in autism spectrum disorders based on animal studies that prove the critical role of oxytocin in the modulation of social cognition. The potential benefits of oxytocin, though, are limited by the biophysical properties of this neuropeptide. This necessitates the development and validation of animal models and behavioral paradigms of social cognition to identify the most efficacious methods of upregulating the oxytocin system. The development of more potent methods stimulating the oxytocin system could have profound effects on social cognitive processes, which could be harnessed clinically. Thus, while oxytocin may play a negligible role in the induction of social impairments of autism spectrum disorders it may still have a meaningful effect on the manifestation of this disorder. It is imperative that the fieldwork fill in the knowledge gaps in the relationship between the oxytocin system and human social cognition, including characterization of the human functional and disorder oxytocin system, the mechanism of action of intranasal oxytocin treatment, and the long-term efficacy of oxytocin administration to realize the clinical potential of oxytocin in the treatment of social impairments [34].
Oxytocin and arginine vasopressin receptors
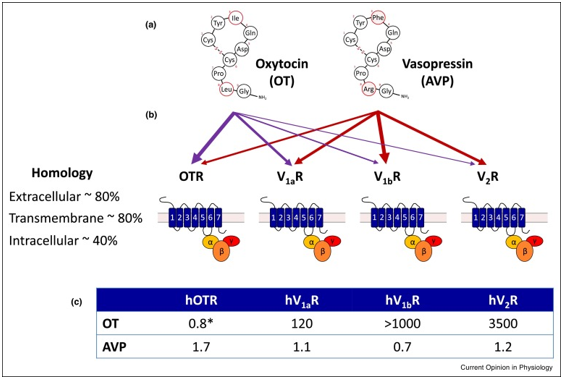
Arginine vasopressin (antidiuretic hormone) is another cyclic neuropeptide released from the posterior lobe of the pituitary gland. Its main functions are to retain water by kidneys and constriction of blood vessels to regulate blood pressure and body fluids volume and osmolarity. It is synthesized by magnocellular neurons in the hypothalamus beside neurons that produce oxytocin neuropeptide. In humans, the structure of oxytocin and vasopressin are very similar, each of them having 9 amino acids and a disulfide bond, and differing only in 2 amino acids (in positions 3 and 8). It also has a short half-life of about 20 minutes and is derived from a preprohormone. Like oxytocin, Arginine vasopressin also signals through G protein-coupled receptors (V1a, V1b, and V2) which have relatively high homology to oxytocin receptors.V1a receptors bind to Gαq/11 proteins and mediate effects via activation of PLC-β whilst V2 receptors bind to Gαs proteins which activate cAMP signaling.V1b receptors can activate several signaling pathways via different G-proteins, according to the concentration of vasopressin and level of receptor expression, e.g. PKC via Gαq/11 and cAMP via Gαs. The similarity in the structure of these two neuropeptides is about 80% homology and the sequence homology of the receptors is high, this characteristic of homology causes cross-reactivity between these peptides and receptors. That is, studies showed that vasopressin binds to its own receptor and to the oxytocin receptors, similarly, oxytocin binds mainly to its receptors and can also, to some extent, activate vasopressin receptors. Oxytocin and vasopressin receptors can also form functional homo- or heterodimers adding further complexity to this system. Vasopressin has been shown to induce contraction of the uterus in humans and other mammals. Uterus in humans has a high sensitivity to vasopressin than to oxytocin. The effect is thought to be mediated via the V1a receptors which, as well as the uterus, are also expressed in the smooth muscle of the blood vessels wall, adrenal cortex, liver, brain, and platelets. The V1b receptors are expressed in the anterior pituitary and adrenal medulla, whilst the V2 receptors are mainly located in the kidneys where it mediates arginine vasopressin antidiuretic action. As with the oxytocin receptors, there is some evidence to suggest that the density of V1a receptors is moderately increased near the end of gestation, and the expression of V1a receptors which suggests the biologically active role of AVP in the pregnant myometrium. The clinical observations that infusion of Arginine vasopressin is able to induce labour (Figure 8) [35].
Oxytocin metabolism, inhibitors, and termination
Oxytocin is administered parenterally and is fully bioavailable. It takes approximately 40 minutes for oxytocin to reach steady-state concentrations in the plasm after parenteral administration. Oxytocin is rapidly removed from the plasma by the liver and kidney. Oxytocinase activity increases throughout pregnancy and peaks in the plasma, placenta, and uterus near term. The placenta is a key source of oxytocin during gestation and produces increasing amounts of the enzyme in response to increasing levels of oxytocin produced by the mother. Oxytocinase activity is also expressed in mammary glands, the heart, the kidney, and the small intestine.Lower levels of activity can be found in the brain, spleen, liver, skeletal muscle, testes, and colon. The level of oxytocin degradation is negligible in non-pregnant women, men, and cord blood. The enzyme oxytocin is largely responsible for the metabolism and regulation of oxytocin levels in pregnancy; only a small percentage of the neurohormone is excreted in the urine unchanged. The plasma half-life of oxytocin ranges from 1-6 minutes. The half-life is decreased in late pregnancy and during lactation [37, 38].
In view of the proposed role of oxytocin in triggering parturition, there has been a long-standing effort to produce specific oxytocin antagonists as a potential means of inhibiting premature uterine contractions. Although several nonapeptide antagonists have been produced and tested, the most-studied oxytocin antagonist to date is the peptide antagonist atosiban, developed by Ferring. Studies attest to the effectiveness of oxytocin antagonists in inducing uterine quiescence in normal labor and in cases of naturally occurring or experimentally induced premature labor. The effectiveness of atosiban to delay premature labor has been demonstrated in many different species, including humans (Figure 9) [39].
It can be concluded that oxytocin is a peptide hormone that is synthesized in brain regions and other organs and posse receptors in many organs. It plays a role in social behavior, emotions love, and affection, the period after childbirth, and metabolic functions. Its potential effects in treating obesity. The half-life time of oxytocin in the brain is triple as long as its half-life time in the periphery. Oxytocin neurons may mediate MC4R-driven sexual behavior. Variations in oxytocin concentration, especially in rapid response to specific experiences. Oxytocin is rapidly removed from the plasma by the liver and kidney. The plasma half-life of oxytocin ranges from 1-6 minutes. The half-life is decreased in late pregnancy and during lactation.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
