
Review Article | DOI: https://doi.org/10.31579/2639-4162/213
1Department of Pathological Physiology Grodno State Medical University, Belarus.
2Department of Pathological Physiology named after D.A. Maslakov, Grodno State Medical University, Belarus.
3Department of Histology, Cytology and Embryology, Grodno State Medical University, Belarus.
45th year student of the faculty of general medicine Grodno State Medical University, Belarus.
*Corresponding Author: Bon E. I, Department of Histology, Cytology and Embryology, Grodno State Medical University, Belarus.
Citation: Maksimovich N. Ye, Bon E. I, Zimatkin S. M, Malykhina A.V, (2024), A New Method for Modeling Total Cerebral Ischemia, J. General Medicine and Clinical Practice, 7(15); DOI:10.31579/2639-4162/213
Copyright: © 2024, Bon E. I. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 09 August 2024 | Accepted: 26 August 2024 | Published: 30 August 2024
Keywords: method for simulating; cerebral ischemia; cerebral cortex
Methodology: The experiments were performed on 12 male outbred albino rats weighing 265±20 g. Total cerebral ischemia in outbred albino rats was modeled by transection of the abdominal aorta. The material was collected at 60 minutes after surgery.
Results: With total cerebral ischemia, a significant decrease in the size of neurons and deformation of the perikarya were observed. Normochromic neurons completely disappeared at the 60th minute. Shriveled neurons made up the majority of cells in the cortical regions studied.
Conclusion: The data obtained on histological changes in neurons of phylogenetically different parts of the cerebral cortex during total cerebral ischemia provide the basis for further detailed study of post-mortem changes in the brain, determining the time of death, creating a fundamental basis for studying the properties of neurons, including their transition from one functional state to another.
Currently, the concept of death in medicine is based on evidence of a stable absence of brain functions. A number of methods are used to diagnose brain function: electroencephalography, assessment of cranial nerve reflexes, and cerebral blood flow studies. In histopathological examination, postmortem changes include edema, hemorrhages, infarctions, necrosis, ischemic softening, shrinkage and deformation of neurons, and pycnosis of their nuclei. In the cerebellar hemispheres, swelling and venous congestion are often detected, and in the subthalamic region and thalamus, areas of spotty lysis are found. The most typical histological change in brain death is considered to be tissue edema with subsequent vascular rupture [8]. Previously conducted studies on the morphological changes in neurons of the parietal cortex and hippocampus in subtotal cerebral ischemia of the brain showed a decrease in the size of perikarya and an increase in the number of hyperchromic and hyperchromic shrunken neurons [3,10]. At the same time, it is of interest to quantitatively study changes in the size, shape and degree of chromatophilia of the cytoplasm of neurons in different periods after total experimental cerebral ischemia. The aim was to analyze changes in the histological characteristics of pyramidal neurons in the parietal cortex and hippocampus of the rat brain during cerebral ischemia, modeled by transection of the abdominal aorta.
The experiments were performed on 12 male outbred white rats with an initial weight of 265±20 g in compliance with the requirements of Directive of the European Parliament and of the Council No. 2010/63/EU of 22.09.2010 on the protection of animals used for scientific purposes. The animals were kept in an air-conditioned room (22 °C) with mixed lighting on a standard vivarium diet and free access to food and water, in groups of no more than 5 individuals per vivarium cage [5].
The use of rats as experimental animals is due to the similarity of the angioarchitecture and morphology of the cerebral cortex in rats and humans [2]. A method for modeling total cerebral ischemia by decapitating animals, leading to its anoxia, is known [4]. However, the existing method has a drawback - the occurrence of respiratory hypoxia due to the transection of the trachea and does not allow for the reproduction of exclusively circulatory hypoxia. In the present study, total cerebral ischemia was achieved by transecting the abdominal aorta in rats under thiopental anesthesia. The material was collected 60 minutes after the operation. After decapitation, the brain was quickly removed, and pieces of the anterior cortex were fixed in Carnoy's fluid. Serial paraffin sections were stained with 0.1% toluidine blue using the Nissl method. The study of histological preparations, their microphotography, morphometry and densitometry of the chromogen sediment in histological preparations were performed using an Axioscop 2 plus microscope (Zeiss, Germany), a digital video camera (LeicaDFC 320, Germany) and the ImageWarp image analysis program (Bitflow, USA). Localization of the parietal cortex and hippocampus in histological preparations of the rat brain was determined using a stereotaxic atlas [13]. In each animal, at least 30 neurons of the fifth layer of the parietal cortex and the pyramidal layer of the CA1 field of the hippocampus were assessed, which ensured a sufficient sample size for subsequent analysis. The number of large pyramidal neurons per unit area of cerebral cortex sections was determined on paraffin sections. Among the total number, cells were isolated by the intensity of cytoplasm staining (chromatophilia). Several types were distinguished: normochromic – moderately colored; hyperchromic – dark; hyperchromic – very dark, with deformed perikarya; hypochromic – lightly colored; shadow cells – almost transparent. The number of each cell type was counted. After preliminary testing for normality of distribution of the indicators, the obtained data were analyzed by nonparametric statistics methods using the Statistica 10.0 for Windows program (StatSoft, Inc., USA). The results are presented as Me (LQ; UQ), where Me is the median, LQ is the value of the lower quartile; UQ is the value of the upper quartile. Differences between the indicators of the control and experimental groups were considered reliable at p<0>
The idea that the degree of intensity of neuronal cytoplasm staining is related to their different functional activity was first expressed by Nissl in 1889. Similar assumptions were made later, but no clear relationship was established. In the 1950s, many works appeared that claimed to solve complex issues of functional morphology. The most widespread point of view was the presence of 3 main groups of nerve cells in the central nervous system: normochromic, hypochromic and hyperchromic, reflecting different functional states of neurons [7,10].
There are opinions that the intense coloration of the neuronal cytoplasm characterizes the predominance of protein formation over its utilization. But there is also information that the hyperchromic neuron, through superexpression of amplified genes, is a cell that intensively synthesizes proteins. Some researchers regard hyperchromic neurons as hyperfunctional and believe that the protein they synthesize is used for their own needs [5].
Shrunken neurons are cells with suppressed functional activity. Their characteristic form is associated with pathological irreversible changes in water-salt metabolism [5,10].
Hypochromic neurons are apparently characterized by an energy-deficient state, and their synthetic activity is low [8].
There is a relationship between the chromatophilia of the cytoplasm of these neurons at the light-optical level and the electron density of the cytoplasm at the electron-microscopic level. The latter is caused by ribosomes, especially free ones, and in hyperchromic wrinkled neurons also by accumulations of a homogeneous osmiophilic substance, probably formed as a result of cell dehydration as a result of a disruption of water-salt metabolism [5].
Hyperchromic shrunken neurons constituted the majority of cells in the studied areas of the cortex at the 60th minute of total cerebral ischemia (Fig. 1, 2, Table 1).
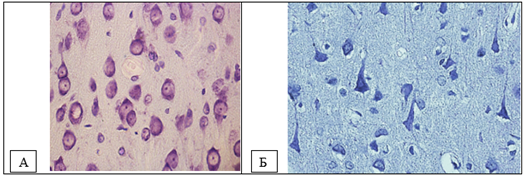
Figure 1. Neurons of the fifth layer of the parietal cortex. A – control (predominance of normochromic neurons), B – total cerebral ischemia (predominance of hyperchromic and hyperchromic shrunken neurons). Digital micrograph. Nissl staining. Magnifying lens x 40.
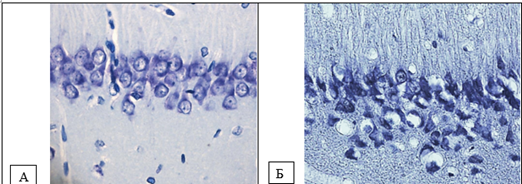
Figure 2. Neurons of the pyramidal layer of the CA1 field of the hippocampus. A – control (predominance of normochromic neurons), B – total cerebral ischemia (predominance of hyperchromic and hyperchromic shrunken neurons). Digital micrograph. Nissl staining. Magnifying lens x 40.
| Index | Control | Total cerebral ischemia | |
| Normochromic neurons/mm2 | 3216(3149;3283) | 67*(0;34) | |
| Hyperchromic neurons/mm2 | 167(134;201) | 100(0;134) | |
| Hyperchromic cellular neurons/mm2 | 33(0;134) | 3618*(3551;3685) | |
| Shadow cells / mm2 | 0(0;134) | 201(134;335) |
Notes
* – p<0>
Table 1 – Number of different types of neurons by the degree of chromatophilia of the cytoplasm of hippocampal neurons in rats with total cerebral ischemia, Me (LQ; UQ)
Hyperchromic neurons are considered as ischemically altered cells [7]. The appearance of wrinkled dark cells in hypoxic and anoxic conditions is a universal and most severe form of reactive and pathological changes in neurons, accompanied by changes in the level of metabolism, tinctorial properties of the cytoplasm, karyoplasm of cells and varying degrees of ultrastructural changes in cytoplasmic organelles. In dark, non-shrunken neurons, RNA synthesis occurs intensively, and pycnomorphic cells contain destructive organelles, their nuclei and cytoplasm become indistinguishable [8]. At the electron microscopic level, compaction of organelles is observed in their cytoplasm. At the same time, the cytoplasm and nucleus of hyperchromic shrunken neurons are reduced in volume, which led to an increase in the density of ribosomes (and, accordingly, ribonucleoproteins) and hyperchromatosis. The number of ribosomes on the outer membrane of the karyolemma is significantly greater than in animals of the control group. A shift of the nucleolus to the periphery of the nucleus and an increase in the concentration of ribonucleoproteins due to their release from the nucleolus and a significant increase in the number of free ribosomes in the cytoplasm of neurons of rats of the experimental group are noted [8]. In shrunken necrotizing neurons, clumps of tigroid substance and neurofibrils usually stick together, and then the cells begin to be diffusely and very intensively stained with thionine and silver [10]. There are opinions that the intense coloration of the neuronal cytoplasm characterizes the prevalence of protein formation over its utilization [12]. But there is also information that a hyperchromic neuron, through superexpression of amplified genes, is a cell that intensively synthesizes proteins. Some researchers regard hyperchromic neurons as hyperfunctional and believe that the protein they synthesize is used for their own needs [5]. Shrunken neurons are cells with suppressed functional activity. Their characteristic form is associated with pathological irreversible changes in water-salt metabolism [5,12].
Depending on the operating conditions, neurons with initial signs of hyper- and hypochromia either transform into shadow cells (hypochromic) or into shrunken hyperchromic neurons with subsequent colliquation and coagulation necrosis or apoptosis [8].
In hyperchromic shrunken neurons, metabolic processes are reduced, the breakdown of nucleoproteins, especially nuclear ones, prevails over their synthesis. The reserves of ribonucleoprotein particles in the nucleus are preserved, but their release into the cytoplasm is blocked.
According to the literature, in the late stages of ischemia, swelling of neurons is observed, accompanied by dissolution of the chromatophilic substance, coarsening, disintegration and melting of neurofibrils, pycnosis of nuclei, thickening and disintegration of processes [6,10]. The neuropil is vacuolated and fragmented, undergoing granular-lumpy disintegration, and myelin dissolves, as a result of which lipid droplets begin to appear along the nerve fibers. Synapses swell, collapse and disappear [10]. The cells decreased in size, becoming more elongated due to deformation of the perikarya [3,11]. At the same time, changes at the 60th minute of total cerebral ischemia reflected deeper destruction of the brain - normochromic neurons were absent, swollen neurons appeared. Hyperchromic neurons were almost never encountered, but wrinkled ones made up the majority of cells in the studied areas of the cerebral cortex. The obtained data on histological changes in neurons of phylogenetically different parts of the cerebral cortex in the dynamics of total cerebral ischemia provide the basis for further detailed study of postmortem changes in the brain, determination of the time of death, creating a fundamental basis for studying the properties of neurons, including their transition from one functional state to another.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
