
Research Article | DOI: https://doi.org/10.31579/2640-1053/067
*Corresponding Author: Sarah Adelaide Crawford, Cancer Biology Research Laboratory, Biology Department, Southern Connecticut State University New Haven, USA.
Citation: Sarah Adelaide Crawford, Alexander Byer-Alcorace and Brielle Hayward-Piakovskyi. Microvesicles Secreted by Glioblastoma Multiforme DBTRG-05MG Tumor Cell Line Contain Proteins Involved in Tumor Invasion, Stemness and Immunosuppression, J. Cancer Research and Cellular Therapeutics, 4(1): DOI: 10.31579/2640-1053/067
Copyright: © 2020 Sarah Adelaide Crawford. This is an open-access article distributed under the terms of The Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 02 March 2020 | Accepted: 19 March 2020 | Published: 30 March 2020
Keywords: microvesicles, glioblastoma, neuroligin-3, integrin-alpha V, beta-tubulin III, insertional membrane editing
Microvesicles or ectosomes are heterogeneous extracellular vesicles ranging in size from 100-1000 nm in diameter. Research suggests that tumor associated microvesicles have a role in immune suppression, multi-drug resistance, invasion, metastasis and angiogenesis via intercellular communication and re-programming. This research study analyzed microvesicles secreted from serum-starved glioblastoma multiforme, the most common brain tumor, noted for having the worst five-year survival rate of any glioma (5%). Live cell imaging studies have shown that microtumor spheroid formation in vitro involves microvesicles that mediate cell-to-cell associations to produce aggregates of cells that coalesce to microtumors, an activity that is consistently observed also in organoid cultures of patient tumors. To study further their role in malignant progression, microvesicles were harvested from the culture medium of serum-starved glioblastoma cell line DBTRG-05MG by ultracentrifugation. Transmission electron microscopy showed that microvesicles comprised the largest fraction of extracellular vesicles, based on measurements of average diameter. Western blot analysis showed that proteins involved in tumor spread, immune system resistance, and early stage neural development were present in microvesicles secreted by the tumor cells. Among these, beta-tubulin III, neuroligin III, and integrin alpha-V were present at levels roughly comparable to glioblastoma whole cell lysates. CD63, CD 47, flotillin-2 and pan-cadherin were also detected in the microvesicle preparations. Notably absent were beta-actin, tsg101, epidermal growth factor receptor and HCAM proteins. A model is proposed, “insertional to mediate tumor invasion and spread.
According to the World Health Organization (WHO), Astrocytoma Grade IV, otherwise known as Glioblastoma Multiforme (GBM), is the most commonly diagnosed adult malignant primary brain tumor, with a 5-year survival rate of only 5.5% [1] and a median survival time estimated between 12 to 14 months from the time of diagnosis [2].Gliomas are glial-precursor or glial cell-derived tumors of the brain, comprised of four main types that include astrocytomas (including GBM), oligodendrogliomas, ependymomas and oligoastrocytomas (mixed gliomas), all of which predominantly arise in white matter [3, 4]. Computerized tomography (CT) and magnetic resonance imaging (MRI) are currently the primary diagnostic tools for GBM [5], which is usually detected at advanced stages of disease when surgery, chemotherapy and radiotherapy are unlikely to result in complete disease remission.
Research suggests that tumor-secreted membrane-bound vesicles budded from the plasma membrane, termed extracellular vesicles (EVs) may potentially serve as biomarkers and/or therapeutic vectors for drug delivery [6] to provide a noninvasive approach to both the diagnosis and treatment of GBM that might improve GBM survival rates. Most, if not all, cells shed EVs into the extracellular environment. EVs are typically divided into three categories based on their size, content and biogenesis [7]. These subsets of EVs include exosomes (EXO), ectosomes or microvesicles (MV), and apoptotic blebs or apoptotic bodies (AB).MVs are large, heterogeneous membrane-enclosed EVs ranging in size from 100-1000 nm in diameter that comprise the bulk of intermediate size-range EVs [8]. Their secretion is the result of a regulated redistribution of membrane lipids followed by the outward budding and fission of the plasma membrane. [9].
Each of the three vesicle types has been shown to communicate intercellularly in various physiological and pathological conditions [10, 11]. Of these, exosomes have been the most studied, while MVs the least. EVs are encased by a lipid bilayer membrane that contains distinct internal cargo, specific to each vesicle type. MVs have a larger diameter than exosomes with a heterogeneous size range from 100-1000 nm in diameter [12]. The third major category of EVs is apoptotic bodies that are released during apoptosis or programmed cell death with diameters ranging from 50-4000nm [12].
The ability of secreted EVs to interact with and transport signaling molecules to other cells and regions of the body presents a new paradigm in our understanding of biological processes in health and disease.To this end, research suggests that EVs are an important mechanistic form of intercellular communication [13]. EVs have been shown to shuttle biological molecules including proteins, lipids, RNA, DNA and metabolites to neighboring cells under normal physiological and pathological conditions [14, 15, 16]. Research on tumor associated microvesicles (TAMVs) suggests that they may play a role in immune suppression, multi-drug resistance, invasion, metastasis, and angiogenesis via intercellular communication and reprogramming [17].
Previous research from our laboratory provided evidence that tumor associated MVs may play an important role in cell-to-cell associations that mediate micro-tumor spheroid formation, an activity that is consistently displayed by tumor cells, both in vitro and in organoid cultures of patient tumors [18]. Live cell imaging studies of glioblastoma microtumor formation in vitro led us to propose that the process of tumor spheroid formation in vitro is characterized by MV mediated cell-to-cell associations leading to spheroid aggregation.
This current study involved the isolation and purification of EVs from serum starved monolayers of the GBM cell line DBTRG-05MG. Electron micrographs of EVs isolated using this production/recovery protocol showed that the isolated the EV fraction is largely comprised of MVs. Western Blot (WB) analysis of MV protein content by immunolabeling demonstrated that MVs produced by this GBM cell line express proteins involved in extracellular matrix (ECM) remodeling, migration, immunosuppression and early stage neurodevelopment. The significance of these observations will be discussed.
Cells and culture conditions
The DBTRG-05MG cell line was obtained from the American Type Culture Collection (ATCC) [catalog number CRL-2020] and cultured in (RPMI) 1640 medium (Thermo Fisher Scientific) ( 1g/L Glucose and 25mM HEPES) supplemented with 5mL 100x Penicillin/Streptomycin (PenStrep) (Fisher Scientific), 5mL 100x Adenine (Sigma), 5mL 100x Hypoxanthine (Sigma), 500uL 1000x Thymidine (Sigma), 5mL 100mM (11mg/ml) sodium pyruvate (Fisher Scientific), 1.25g Glucose (Sigma), 1g sodium bicarbonate and 10
Isolation and purification of extracellular vesicles from serum-starved DBTRG-05MG glioblastoma monolayers
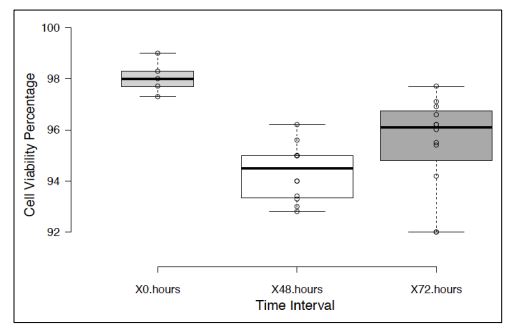
Monolayer cultures of DBTRG-05MG GBM cell line at approximately 75% confluence were incubated in serum-free RPMI for 48-72 hours to stimulate EV production, followed by differential ultracentrifugation to pellet cell fraction enriched with (MVs) (see Materials and Methods). Trypan blue viability assays of monolayer cells after cultivation in serum-free medium showed approximately 95% cell viability (Fig. 1), indicating that the vesicles produced by this method did not consist of a significant fraction of apoptotic bodies.
Transmission electron microscopy (TEM) of extracellular vesicles isolated by differential centrifugation
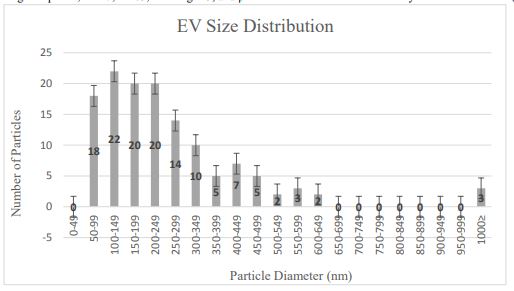
Size data collected from TEM images of EVs prepared from serum-starved cells were used to determine the overall EV morphology and particle distribution in the sample (Fig. 2).

TEM particle measurements taken for approximately 130 vesicles showed that the majority (75%) of the vesicles ranged in diameter from 100-500 nm, in the size range characteristic of MVs. Approximately 15% EVs showed a diameter of less than 100 nm, suggesting they may be comprised of exosomes of endosomal origin. The EM data showed that the EV isolation protocol allowed the recovery of EVs predominantly in the size range of MVs.
Western blot analysis of MV protein composition
MV lysates were analyzed against a panel of primary antibody probes and compared to whole cell lysates (WCLs) of DBTRG-05MG cells from which the MVs were isolated.

Each of the proteins tested (Table 1) was present at detectable levels in WCLs prepared from DBTGR-05MG monolayer cultures. Among these, pan-cadherin, flotillin-2, integrin alpha V, CD47, CD63, neuroligin-3, and β-tubulin-III were also detected in lysates of GBM MVs (Fig. 3).
Although the relative amounts of these proteins in MVs as compared to whole cell lysates were difficult to quantitate, OD600 spectrophotometry measurements of MV lysates as compared to whole cell lysates indicated that MV density was approximately one-third that of whole cell lysates used in Western blot analyses (data not shown). The intensity of the bands corresponding to integrin alpha V, neuroligin-3 and β-tubulin III were comparable in MV and WCLs, despite the fact that the total protein content in the WCL was significantly greater. In contrast, the relative levels of CD63, CD47, pan-cadherin and flotillin-2 appeared to be significantly lower in MVs than WCLs, based on staining intensity. Several proteins present at detectable levels in WCLs were not observed in lysates prepared from MVs produced by these cells. Notably absent from MV lysates were B-actin, Tsg101, HCAM and EGFR (Fig. 4).

Research suggests that EVs mediate communication within the tumor microenvironment, contributing to cancer progression [21] and metastasis [22, 23]. The research reported in this paper demonstrates that some proteins linked to GBM invasion and spread are also present in tumor-secreted microvesicles (Table 1). High levels of integrin αV, a transmembrane ECM adhesion protein, were detected in GBM EVs. Biopsies from five cerebral GBM patients were shown by Gladson and Cheresh [24] to express vitronectin and the αV-β3 integrin receptor, which was not found in low grade astroglial tumors, reactive astrogliosis or normal cortex or cerebral white matter. Moreover, cultured GBM cells were shown to require vitronectin receptor for brain tissue attachment and, therefore, could not attach to normal brain tissue. This mode of adhesion may be a requirement for GBM invasion into normal brain tissue, potentially implicating GBM secreted MVs containing integrin αV in this process. Francovic et al. [25] have demonstrated that integrin αV upregulation is correlated with tumor progression from low grade to aggressive gliomas, directly implicating this receptor complex as a biomarker for invasion and metastasis. The study authors suggested that integrin αV may be involved in escape from senescence.
Β-tubulin III, a biomarker for tumor aggressiveness and resistance to taxanes, is a poor prognosis indicator whose upregulation is induced by oxidative stress [26]. Its presence in tumor secreted MVs suggests that diagnostic screening of cerebral spinal fluid for MVs containing this biomarker could be a valuable addition to current assessment protocols.
Very high levels of neuroligin-3 and its 75kDa cleaved product were also detected in MVs from GBM tumor cells. Neuroligin-3 activates multiple tumor promoting signal pathways that enhance cell proliferation. An important mechanism that mediate neural regulation of brain cancer is activity-dependent cleavage and secretion of this synaptic adhesion molecule, which promotes glioma proliferation through the PI3K-mTOR pathway [27]. Research has shown that when this signaling pathway is blocked, high grade gliomas cannot proliferate in mice [28]
CD47 blocks phagocytosis of tumor cells by binding to SIRPα receptors on microglial phagocytes; this represents an important component of immune system evasion by tumor cells [29]. Likewise, MVs expressing CD47 secreted by GBM tumor cells may play a direct role in immunosuppression by blocking microglia associated tumor cell phagocytosis. To this end, therapeutic inhibition of CD-47/SIRP-alpha by humanized antibodies has been developed as a treatment for pediatric brain tumors as it activates microglial phagocytosis of brain tumor tissue [30].
CD63 is a tetraspanin that plays an important role in cell adhesion and motility; it is a biomarker for poor prognosis when detected in many types of cancer. [31]. Interestingly, this protein is expressed in undifferentiated neural stem cells, but not in their differentiated counterparts [32]. Its presence in GBM MVs further implicates these vesicles in activities associated with tumor stemness and progression. CD44 is a previously identified biomarker associated with tumor migration and adhesion to the extracellular matrix via binding to hyaluronic acid [33]. Pan-cadherin was also detected in GBM MVs; cadherins are transmembrane epithelial cell-to-cell adhesion proteins regulated via calcium-mediated interactions. Additional research by Kaur et al. [34] showed that cadherin-11 is expressed in human glioma cell lines and is associated with tumor cell migration and growth factor independence.
Surprisingly, B-actin was not detected in MVs secreted from GBM cells. It is possible that MV budding from the plasma membrane may occur in actin-poor regions in the cell membrane where the absence of cytoskeletal components may facilitate membrane extrusion to form extracellular vesicles. In contrast, the presence of flotillin-2 in secreted MVs suggests that cell membrane areas containing this membrane protein may be associated with MV budding and release. The absence of detectable levels of tsg101, a biomarker for exosomes, is consistent with our data showing that the EV preparation used in this study consisted largely of EVs in the size range characteristic of microvesicles (MVs).
Cultivation of monolayer GBM cells in serum-free medium for 48-72 hours enhanced MV production without significant loss of culture viability and associated production of apoptotic bodies. Moreover, the ultracentrifugation protocol used in this study to recover MVs produced by serum-starved GBM cells permitted a high yield isolation of EVs in the particle size range of MVs, based on TEM size range data. A small, but significant (15%), fraction of the recovered EVs were in the size range (50-100nm) of exosomes, and this may have accounted for the cup-shaped morphology observed for some of the vesicles in TEM imaging studies.
An important question raised by this and similar studies is the connection between serum depletion and increased EV production by cultured cells. It is probable that serum starvation affects cell-to-cell and cell-to-substrate adhesion parameters in cultured cells to promote EV production. For example, the major source of fibronectin in vitro is serum. As a consequence of serum starvation, decreases in the levels of proteins important in cellular attachment may act as a trigger for MV production; moreover, cell detachment may represent an important trigger for EV formation in both physiological and in vitro conditions, consistent with their suggested role in cell movement and migration. It is possible that the composition of secreted MVs may vary depending on the conditions in which their production is induced; therefore, it will be important to characterize the protein composition of MVs induced by physiological factors as well. Previous research in our laboratory has shown that MV release from cultured GBM cell line DBTRG-05MG is stimulated by CaCl2 [18]. Unlike serum starvation, CaCl2 mediated induction of MV production represents a physiologically relevant condition. In cancer patients, tumor induced hypercalcemia, a poor prognostic parameter, may contribute to tumor progression, in part, by contributing to a tumor microenvironment that promotes tumor spread and metastasis associated with and related to elevated MV production. Despite the fact that CaCl2 represents a more physiological trigger of MV production, serum starvation produces a higher, more consistent yield of tumor MVs without inducing significant apoptosis.
That cell membrane biomarkers of GBM tumor invasion and progression are also displayed in tumor secreted MVs supports the role of these vesicles in these processes. The secretion of MVs containing proteins involved in tumor invasion/spread, immunosuppression and “stemness” may represent an important mechanism of glioma progression. In this context, MVs may serve as an intricate intercellular communication platform for initiating invasion and tumorigenesis within the tumor microenvironment, leading to tumor progression and metastasis. Research suggests that the overriding function of EVs is the transport and deposition of cargo from the cell of origin to a cellular recipient and to exert a primary effect on cell biology through this mechanism [35]. MMVs may, however, exert a more comprehensive and fundamental role in tumor biology by modifying the tumor niche to induce stromal malignant transformation and tumor spread by morphogenetic remodeling of the tumor microenvironment. The proposed model by which this process occurs is termed “insertional membrane editing”. (Fig. 5).

This model suggests that MVs secreted by tumor cells bind to the membranes of stromal cells and, in so doing, insert membrane components from the original tumor cell into the recipient cell membrane that induce stromal tissue transformation. Secreted MVs are mobile elements whose production may be initiated by the disruption of tissue architecture caused by primary tumor hyperplasia. Subsequent movement about the stromal niche lead to interactions with stromal cells to induce tissue morphogenesis by membrane insertion. The result is an “edited” stromal cell membrane that contains proteins that may alter the gene expression profile of the recipient cell via the induction of signal transduction cascades that reprogram the gene expression to elicit changes in the stromal recipient associated with malignant transformation. Tumor invasion into normal regions of the brain may, therefore, be initiated, in part, by the uptake of MVs shed from tumor cells. In the tumor microenvironment, changes in cell-to-cell adhesion parameters, such as those involved in epithelial-to-mesenchymal transitions (EMTs), may further promote the production of EVs to amplify tumor invasion and spread, ultimately remodeling the stromal tissue into a tumor invasion zone, associated with continuously amplifying signals for tumor cell migration, ECM attachment and invasion. In this context, insertional membrane editing may be a fundamental component of tumor progression by which tumor associated MVs educate the stromal niche to express critical parameters of malignant transformation that contribute to advanced, systemic disease. By this mechanism, a regionally localized tumor may dispense migratory carriers of key inducers of malignancy, allowing its effects to extend beyond its primary location to effect local microenvironmental and, ultimately, systemic changes that characterize advanced high-grade malignancy.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
