
Research Article | DOI: https://doi.org/10.31579/2643-1912/002
1 Deportment of Pharmaceutics, Jangaon Institute of Pharmaceutical Sciences, India.
*Corresponding Author: Stalin Reddy Challa, Deportment of Pharmaceutics, Jangaon Institute of Pharmaceutical Sciences, India.
Citation: Stalin Reddy Challa and Swathi Goli, Differentiation of Human Embryonic Stem Cells into Engrafting Myogenic Precursor Cells. J. Stem cell Research and Therapeutics International, DOI: 10.31579/2643-1912/002
Copyright: © 2019 Stalin Reddy Challa, This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 16 March 2019 | Accepted: 09 April 2019 | Published: 11 April 2019
Keywords: muscle tissue; human embryonic stem cells; myogenic precursor; immunodeficient
Degenerative muscle diseases affect muscle tissue integrity and function. Human embryonic stem cells (hESC) are an attractive source of cells to use in regenerative therapies due to their unlimited capacity to divide and ability to specialize into a wide variety of cell types. A practical way to derive therapeutic myogenic stem cells from hESC is lacking. In this study, we demonstrate the development of two serum-free conditions to direct the differentiation of hESC towards a myogenic precursor state. Using TGFß and PI3Kinase inhibitors in combination with bFGF we showed that one week of differentiation is sufficient for hESC to specialize into PAX3+/PAX7+ myogenic precursor cells. These cells also possess the capacity to further differentiate in vitro into more specialized myogenic cells that express MYOD, Myogenin, Desmin and MYHC, and showed engraftment in vivo upon transplantation in immunodeficient mice. Ex vivo myomechanical studies of dystrophic mouse hindlimb muscle showed functional improvement one month post-transplantation. In summary, this study describes a promising system to derive engrafting muscle precursor cells solely using chemical substances in serum-free conditions and without genetic manipulation.
Muscular dystrophies are debilitating degenerative muscle diseases caused by defective muscle proteins that disrupt normal homeostasis [1,2]. During embryogenesis Pax3 and Pax7-expressing cells drive muscle formation, growth and repair [3-8]. In the postnatal period, these cells are termed satellite cells –stem cells that remain quiescent beneath the basal lamina of muscle fibers under normal conditions. Tissue damage triggers these cells to proliferate and differentiate into myoblasts, leading to the formation of multinucleated myotubes and fibers [9-12]. However, dystrophic tissue induces repeated activation of satellite cells, ultimately leading to stem cell exhaustion, limiting the ability for further restorative myogenesis. Given their role in muscle formation, growth and repair, satellite cells, or other myogenic precursors with similar characteristics, would be attractive candidates for cell-based therapies for muscular dystrophies. Successful cell therapy would require myogenic precursor cells to: i. repopulate damaged muscle, ii. contribute to the formation of muscle fibers, and iii. lead to expression of normal muscle proteins in host tissue [13]. In addition, an important therapeutic benefit would be repopulation of the muscle stem cell niche to allow for ongoing homeostasis.
Embryonic stem cells (ESC) have the capacity to expand indefinitely, while maintaining the ability to differentiate into a wide variety of cell types. As such ESC may be useful for cell transplantation therapy or tissue engineering. However, a non-genetics method to obtain hESC derivatives with muscle regenerative abilities is lacking. Promising attempts to derive human myogenic precursors from pluripotent stem cells have relied on viral–mediated MYOD-expression [14-18], serum-containing media [14,15], use of demethylating agents [15], or selective multi-step cell culture approaches and cell sorting procedures that may be difficult to implement on a large-scale [16]. The most efficient methods described to date used genetic manipulation of human ESC (hESC) to overexpress the myogenic transcription factors, PAX3 and PAX7 [19]. Importantly, the use of genetically manipulated or virally induced cells, as well as cells exposed to serum that contains non-human components, may be difficult to use in human trials due to safety concerns.
Therefore, in order to establish alternative ways of driving early myogenic specification, we leveraged previous studies describing chemical compounds that stimulate and facilitate hESC differentiation toward paraxial mesoderm, the lineage responsible for limb and limb-associated skeletal muscle development. hESC can be chemically stimulated by activation of the bone morphogenetic protein (BMP) pathway, which is a key inducer of mesoderm formation during embryogenesis [14,20-24]. Treatment with the phosphatidylinositol 3-kinase (PI3K) inhibitor, Ly294002 hydrochloride (Ly), facilitates the differentiation of hESC by reducing self-renewal ability [25,26]. Additionally, paraxial mesoderm, the lineage responsible for limb muscle development, can be formed by brief stimulation with BMP and inhibition of the TGFb/Activin pathway using the small molecule, SB431542 (SB) [27]. This compound specifically inhibits the Activin pathway without affecting BMP signaling. It has been shown that inhibition of the TGFb/Activin pathway using SB up-regulates myogenic genes during hESC differentiation in defined culture conditions [14]. Another important factor in mesoderm formation is fibroblast growth factor (FGF), which also determines myogenic fate during development and plays a role in satellite cell activation [28-31]. Therefore, it is plausible that these components could be used to drive myogenesis in hESC cultures.
Based on these observations, we hypothesized that engrafting myogenic precursors could be derived from hESC by the sequential and coordinated manipulation of the BMP, TGFß and FGF pathways. Importantly, we sought to prove that myogenic specification of hESC could be achieved without the use of allogeneic components in serum or genomic alteration. To this end, we tested whether myogenesis could be induced through embryoid body (EB) differentiation in medium containing basic fibroblast growth factor (bFGF), Ly and low levels of FGF, in combination with short exposure to BMP4 (referred to as KFLy(B)S medium). Separately, a combination of FGF and SB alone was tested for the ability to drive myogenesis (referred to as KFS medium). We determined the capacity of hESC-derived myogenic precursors to engraft in skeletal muscle upon transplantation in hindlimb muscles of SCID mice. In addition, we determined functional changes in transplanted muscle of mdx mice using ex vivo myomechanical analysis. The novel differentiation approach described in this study circumvents many of the limitations encountered in the transition of pluripotent stem cells into myogenic precursor cells, while enhancing our understanding of the mechanisms that govern stem cell differentiation and skeletal myoge
Human ESC culture and myogenic differentiation
All work with hESC was done with the approval of UCSF Stem Cell Research Oversight Committee. The H9 hESC (WA09; WiCell) were maintained on irradiated CF1 mouse embryonic fibroblasts (MEF; Millipore, Temecula, CA) in KSR medium [Knockout Dulbecco’s Modified Eagle Medium containing 20% Knockout Serum Replacement, 2mM glutamine, 0.1 mM nonessential amino acids, 0.1 mM β-mercaptoethanol (all from Invitrogen, Carlsbad, CA)] supplemented with 12.5 ng/ml recombinant human fibroblast growth factor basic (bFGF; R&D Systems, Minneapolis, MN) as previously described [32].
hESC colonies were detached after brief exposure to Collagenase IV (1 mg/ml; Sigma-Aldrich, St. Lois, MO), washed and collected by settlement. Differentiation was initiated by embryoid body (EB) formation in pre-differentiation medium KFLy(B)S: KSR medium freshly supplemented with 20 ng/ml bFGF, 10 μM Ly294,002 hydrochloride (Ly; Sigma-Aldrich), 10 μM SB431542 hydrate (SB; Sigma-Aldrich), and for the first 36 hours supplementing with recombinant human bone morphogenetic protein-4 (BMP-4) (10 ng/ml; HumanZyme, Chicago, IL), or pre-differentiation medium KFS: KSR medium freshly supplemented with 40 ng/ml bFGF and 10 μM SB. Both pre-differentiation media were supplemented one time with 10 μM Rho-associated protein kinase inhibitor (ROCKi) at the day of EB formation. SB and Ly were reconstituted in dimethylsulfoxide (DMSO) at 20 mM, and diluted to the final concentration in pre-differentiation medium. Human EBs were kept in suspension in ultra-low cluster 6-well plates (Costar, Corning, NY) and placed onto an orbital shaker (Daigger, Vernon Hills, IL) at a speed of 90 rpm. After 36 hours, medium was refreshed, which was repeated every other day until day six.
To assess the myocyte-forming potential of EB-derived myogenic precursor cells derived in KFLy(B)S and KFS (differentiation day 6) media, EBs were plated onto 12.5 μg/cm2 fibronectin (from human plasma; Sigma-Aldrich) in 0.1% gelatin-coated culture surfaces in pre-differentiation medium to stimulate cell outgrowth. After two days, medium was replaced with muscle differentiation medium [DMEM-high glucose, 10% fetal bovine serum (FBS; Hyclone, Thermo Scientific, South Logan, UT), 5% horse serum (HS; also from Hyclone) supplemented with 5 ng/ml bFGF] and cultured for an additional 4 days.
Quantitative real-time PCR
RNA was extracted from cells using the RNeasy Mini Kit including DNase treatment (Qiagen, Valencia, CA) according to the manufacturer's instruction. RNA concentration was measured using a Nanodrop ND-1000 Spectrophotometer (Nanodrop Technologies) to convert equal quantities of mRNA into cDNA using the Superscript™ III First-Strand Synthesis System (Invitrogen). Relative gene expression was determined using TaqMan Assay (Applied Biosytems) on an ABI 7300 Real-Time PCR system with the following human primer pairs (ABI): OCT4 (Hs00742896_s1); Brachyury (T) (HS00610080_m1); PAX3 (Hs00240950_m1); PAX7 (Hs00242962m1); MYOD (Hs00159528_m1); Desmin (Hs01090875_m1); Myogenin (Hs002311167_m1); MYHC (Hs00430042_m1) and GAPDH (4326317E). Cycle threshold (Ct) value for detection of gene of interest was normalized against Ct value of the housekeeping gene, human GAPDH, and relative changes were calculated according the ΔΔCt method. Results of relative fold expression were analyzed using GraphPad prism software in a two-way ANOVA with Bonferroni post-test.
Immunostaining
Immunofluorescence staining was performed on EB-outgrowths cultured in of KFLy(B)S or KFS with mouse anti-human Pax7 (undiluted), Myogenin (F5D; undiluted) and MyHC (MF20; 1:20) antibodies (all three supernatants from Developmental Studies Hybridoma Bank); mouse anti-human MyoD (5.8A; 20 μg/ml; R&D systems); or mouse anti-human Desmin (D33; 10 μg/ml; Dako, Glostrup, Denmark). Cells were fixed in 4% paraformaldehyde (PFA; EMS, Fort Washington, PA) for 15 min. and permeabilized in 0.4% Triton-X100 for 10 min. After blocking for 30 min in blocking solution [2% normal goat serum (NGS; Dako), 1% bovine serum albumin (BSA; Sigma-Aldrich), 0.1% cold fish gelatin (Sigma-Aldrich), 0.1% Triton-X100, 0.05% Tween-20 in phosphate buffered saline (PBS)], cells were subsequently incubated with the primary antibodies diluted in blocking solution for 1 hr. After washing (three times; 5 min in PBS with 0.05% Tween-20), primary antibodies were detected using Alexa Fluor® 488-labeled goat anti-mouse (1:200; Life Technologies, Grand Island, NY) for 30 min. All steps were performed at room temperature (RT). Fluorescence microscopy was performed using a Nikon Eclipse TE300 microscope and Zeiss AxioScope Imager Z.2 with Apotome (Zeiss, Inc, Thornwood, NY).
Intramuscular cell transplantation
All experiments involving animals were done with the approval of the UCSF Institutional Animal Care and Use Committee. Hindlimbs of CB17.B6-PrkdcscidLyst bg/Crl (8-12 wk-old) and C57BL/10ScSn-Dmdmdx/J (X-linked muscular dystrophy; aged 6-8 weeks) mice (Jackson Laboratories, Bar Harbor, ME) were exposed to gamma radiation (5 Gy and 18 Gy, respectively) three days before cell transplantation to inhibit muscle regeneration by host muscle cells. Mice cages were supplied with a water bottle containing an antibacterial oral suspension (200 mg sulfamethoxazole-40 mg trimethoprim/ 200 ml water) from the day of irradiation through euthanasia. For intramuscular injection, EB-derived in KFLy(B)S or KFS were trypsinized and filtered through 40 μm nylon cell strainer (Gibco) to obtain a single cell suspension. One million cells cultured in KFLy(B)S or KFS were suspended in 50 μl PBS (supplemented with 10 μM ROCKi) and injected into the tibialis anterior (TA) muscle at two sites. As control, the contra-lateral muscle was injected with 50 μl PBS with 10 μM ROCKi. TA muscles were analyzed by immunohistochemistry one month post-transplantation, as described above.
Immunohistochemistry
TA muscles were embedded in OCT, snap-frozen in liquid nitrogen and stored at -80°C. Cryosections (5 mm) were fixed in acetone (ice cold, 10 min) and blocked in blocking solution [5% NGS, 5% Normal Donkey Serum (Dako), 5% BSA, 0.1% cold fish gelatin, 0.1% Triton-X100, 0.05% Tween-20 in PBS]. Sections were incubated with rabbit anti-human Dystrophin (1:2500; Novus Biologicals, Littleton, CO), chicken anti-mouse Laminin (2 mg/ml; Abcam), mouse anti-human Desmin (10 μg/ml; D33; Dako), or mouse anti-human HLA-1 (W6/32; antigen determinant common to HLA-A, B and C; 4 mg/ml; provided by Frances Brodsky, Dept. of Biopharmaceutical Sciences, UCSF) in blocking buffer for 1 hour. Alexa488/594-conjugated goat anti-mouse IgG, Alexa594-Donkey anti-Rabbit IgG, or Alexa488-Goat anti-Chicken IgG (1:250; all from Life Technologies) were used to detect primary antibodies. Sections were then washed and dehydrated in ethanol (70%, 90%, 100%; 5 min each), air-dried, and mounted in Vectashield with DAPI (Vector, Burlingame, CA). For immunohistochemical detection, sections were additionally blocked for endogenous peroxidase (0.3% H2O2 in PBS, 20min). Primary antibody HLA-1 was detected by peroxidase labeled goat anti-mouse. Visualization was performed by a 3-amino-9-ethyl-carbazole (AEC; Sigma) substrate incubation for 4 minutes. Nuclei were counterstained with Mayer’s haematoxyline (2 min, Fluka, Buchs, Zwitserland). All steps were performed at RT. Fluorescence microscopy was performed using a Nikon Eclipse TE300 microscope and Zeiss AxioScope Imager Z.2 with Apotome (Zeiss, Inc, Thornwood, NY).
Ex vivo myomechanical analysis of transplanted EDL muscle in mdx mouse model
Ex vivo myography was performed to determine the increase in force production and to determine muscle fatigue level after cell transplantation into the extensor digitorum longus (EDL) muscle of mdx mouse. Myomechanical analysis was performed as previously described [33,34]. Force-frequency fatigue was measured by exposing muscle to a supramaximal stimulus train of 3-5 pulses (300 msec duration separated by 3 sec) at successive frequencies (30, 60, 100 and 140 Hz), with 5 minute intervals between stimulations. Low-frequency time-fatigue was measured by supramaximal tetanic muscle stimulation at low frequency (60 Hz) for duration of 300 msec, repeated every 3 seconds for a period of 10 minutes. For myo-mechanical analysis, the left and right leg variables of an animal were averaged in each group and muscle mass, cross-sectional area, and length was measured. Significant differences were present between the means of each group compared to the PBS group. Multiple t-tests were performed with the number of comparisons adjusted by Bonferroni correction. A value of p <0.05 was considered significant.
nhibition of TGFβ pathway directs myogenic commitment
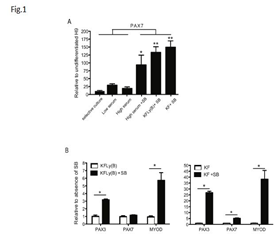
To compare previously described, serum-containing methods for their capacity to induce the formation of myogenic precursors, we differentiated hESC by a selective culture approach using serum-containing (10% FBS) medium to enrich for CD73+ mesodermal stem cells followed by a second cell sort for expression of neural cell adhesion molecule (NCAM), a molecule also expressed by myogenic cells [16]. Furthermore, we differentiated hESC-EB in low serum conditions (2% HS) [35-37] and high serum conditions (10% FBS) during 2-4 weeks in culture [15]. We observed expression of PAX7 in cells derived through the selective multi-step methods (preCD73/NCAM sort) and when exposed to low and high serum conditions (Figure 1A). Addition of SB, a compound that inhibits the TGFb pathway, to cells differentiated in 10% FBS culture conditions resulted in markedly increased PAX7 levels (Figure 1A) *p<0.01. This indicated the myogenic stimulatory effect of TGFb inhibition in hESC. Next, we transiently stimulated the BMP pathway with BMP4 during EB differentiation combined with Ly and high levels of bFGF in serum-free conditions (KFLy(B)). This resulted in high levels of PAX7 expression (Figure 1A) p<0.001. This was also seen when EB were exposed solely to high concentrations of FGF (KF) (Figure 1A) p<0.001.
Myogenic differentiation in EB culture
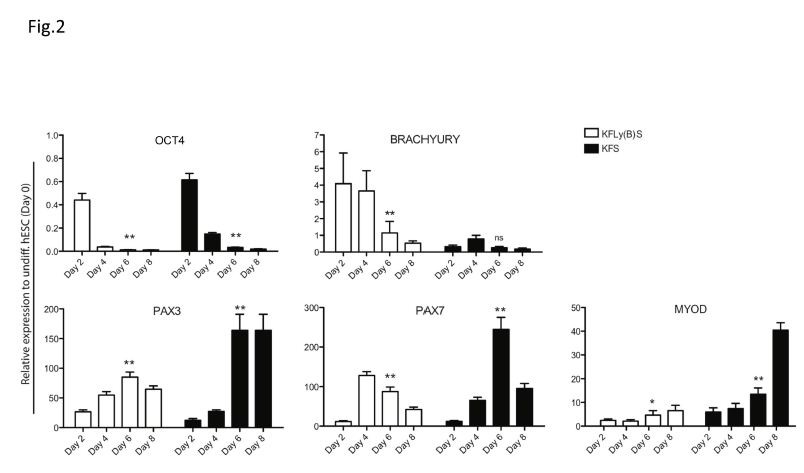
We hypothesized that myogenic precursor cells would be more likely to engraft in host muscle, and have a greater chance of repopulating the stem cell niche, than fully differentiated myoblasts. To monitor the stages of myogenic differentiation, we cultured hESC-EB in serum-free KFLy(B)S or KFS and examined myogenic gene expression by qRT-PCR at days 2, 4, 6 and 8, relative to undifferentiated hESC (day 0). OCT4 expression was used as an indicator of residual pluripotency after initiation of hESC differentiation. In both differentiation conditions, OCT4 expression fell dramatically by day 2 and further diminished through day 6 of differentiation (Figure 2) p<0.001.
KFLy(B)S- and KFS-derived precursors differentiate into myoblasts in vitro
Directed differentiation of stem cells can be stimulated with exposure to specific extracellular matrix (ECM) components they would likely encounter in the stem cell niche [42]. This has specifically been shown for stem cell differentiation toward the myogenic lineage [43,44]. EB differentiated for 6 days in KFLy(B)S or KFS were plated in dishes coated with Matrigel, laminin or fibronectin to stimulate myogenic cell outgrowth. We observed that attachment to fibronectin-coated dishes, compared to Matrigel and laminin, more effectively promoted myogenic differentiation (data not shown). Therefore, fibronectin coating was used to evaluate the myogenic potential of EB-outgrowth cells (EB-O) during the subsequent 6 days in muscle differentiation medium.
PAX3 and PAX7 are critical factors in embryonic skeletal myogenesis and participate after birth in muscle homeostasis. They are, therefore, important indicators of a myogenic precursor stage that harbors regenerative capacity. Several methods have been published describing the myogenic differentiation of hESC, however, limitations of published methods include the use of non-human serum and genetic manipulation that may hamper therapeutic application due to safety concerns [48,49]. This study underscores that myogenesis can be achieved in hESC cultures in a short time frame when exposed to appropriate developmental cues. We have demonstrated two approaches to derive PAX3- and PAX7-expressing myogenic precursors from hESC using a novel sequence of chemical compounds that direct a myogenic cell fate. Upon transplantation, these muscle precursor cells engrafted into muscle, formed new fibers, and improved muscle strength.
In the course of this study a publication revealed the success of a chemically defined culture system in transgenic zebrafish that directs skeletal myogenesis of human induced pluripotent stem cells [50]. Therefore, the combination of pathway inhibitors and stimulators described in this study provide an effective platform to advances approaches to muscle differentiation from hESC. Among the benefits are the avoidance of the use of serum and genetic alteration to promote gene expression. Furthermore, we showed that hESC entered the myogenic program in less then one-week making this approach faster than other published methods applying hESC differentiation during 2-4 weeks. Additionally, lacking selective cell culture approaches contributed to a method that is costs effective and easy.
In mdx mice, transplantation with hESC-derived precursors derived using these methods resulted in improved mechanical force generation and decreased fatigue. Both muscles injected with KFLy(B)S and KFS-derived cells demonstrated higher maximal tension than muscle injected with PBS or EDL that was not injected with cells, indicating improved functional capacity. Interestingly, EDL from mice injected with KFLy(B)S-derived cells demonstrated greater maximal tetanic tension, compared to both EDL from mice injected with PBS and KFS-derived cells and EDL that was not injected. However, force-frequency analysis demonstrated decreased fatigue in muscle from mice injected with KFS-derived cells compared to the other groups. The mechanisms that might account for these differences are unclear, but it is intriguing that the ESC-derived precursors could alter functional capacity by affecting different muscle properties (e.g strength vs endurance). Most importantly, however, the findings of the myomechanical analysis indicate preliminary proof-of-concept for functional myogenic potential for these hESC-derived precursor cells.
We have shown that a short exposure of hESC to chemically defined media promotes differentiation of hESC into engrafting myogenic precursor cells that can contribute to the formation of new fibers in vivo and improve muscle strength as analyzed ex vivo. These techniques are less laborious than previously published methods, and the avoidance of xenobiotic agents and serum may mitigate safety concerns. Further studies to assess durability of transplanted cells and newly formed fibers, as well as their contribution to muscle homeostasis over time are warranted.
Dear Editorial Team, Clinical Medical Reviews and Reports. My experience with the journal was highly positive. The peer-review process was rigorous, constructive, and completed in a timely manner. The reviewers provided valuable comments that helped improve the quality and clarity of our manuscript. The editorial office was professional, responsive, and supportive throughout all stages of the publication process. Communication was clear and efficient, and any questions were addressed promptly. Overall, I found the journal to maintain high scientific standards and an excellent publication workflow. I would be pleased to consider submitting future work to this journal. Best wishes from, Elena Popa.

It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,
